
Статья 9 ТК РФ с комментариями
Полный текст ст. 9 ТК РФ с комментариями. Новая действующая редакция с дополнениями на 2021 год. Консультации юристов по статье 9 ТК РФ.
В соответствии с трудовым законодательством регулирование трудовых отношений и иных непосредственно связанных с ними отношений может осуществляться путем заключения, изменения, дополнения работниками и работодателями коллективных договоров, соглашений, трудовых договоров.
Коллективные договоры, соглашения, трудовые договоры не могут содержать условий, ограничивающих права или снижающих уровень гарантий работников по сравнению с установленными трудовым законодательством и иными нормативными правовыми актами, содержащими нормы трудового права. Если такие условия включены в коллективный договор, соглашение или трудовой договор, то они не подлежат применению .
Комментарий к статье 9 ТК РФ
1. В комментируемой статье реализован один из основополагающих методов отрасли трудового права — сочетание нормативного и договорного регулирования трудовых отношений.
Истоки коллективно-договорного регулирования трудовых отношений можно найти в Англии, а позднее во всей Европе. В России «коллективные договоры появились в самом начале 20 века, но правовое закрепление получили лишь в Кодексе законов о труде 1918 года и Положении о порядке утверждения коллективных договоров 1918 года».
________________
Стрижаков Г. Правовая природа коллективного договора как акта социального партнерства // Трудовое право. 2007. N 10.
Сегодня коллективные договоры и соглашения являются одними из важнейших источников трудового права, а роль договорного регулирования трудовых и непосредственно связанных с ними отношений возрастает.
Договоры и соглашения о труде заключаются на различных уровнях социального партнерства, соответственно выполняют разную роль при регулировании отношений в сфере труда. Если коллективный договор регулирует социально-трудовые отношения в организации или у индивидуального предпринимателя, то коллективные соглашения на федеральном, межрегиональном, региональном, отраслевом (межотраслевом) и территориальном уровнях социального партнерства.
Ю.П.Орловский отмечает, что договорное регулирование должно осуществляться в соответствии с трудовым законодательством, то есть соответствовать ему, развивать и конкретизировать его положения.
________________
Комментарий к Трудовому кодексу Российской Федерации / Ответственный редактор Ю.П.Орловский. М.: Контракт, Инфра-М, 2002. С.29.
Закрепляя содержание коллективного договора, соглашений и трудового договора (ст. 41, 46 и 57 соответственно), законодатель, по сути, определяет круг вопросов, которые, как правило, регулируются указанными соглашениями о труде. Однако допускается регулирование в договорном порядке и других отношений, при условии, что ТК РФ не устанавливает исключительное централизованное регулирование данных отношений.
В большинстве случаев ТК РФ прямо предусматривает регулирование тех или иных отношений коллективными договорами и соглашениями или трудовым договором. При этом может быть указание как на отдельные (или коллективный договор, или соглашение, или трудовой договор), так и на все виды договоров о труде.
Например, в соответствии с ч.2 ст. 270 ТК РФ для работников в возрасте до восемнадцати лет, поступающих на работу после окончания общеобразовательных учреждений и образовательных учреждений начального профессионального образования, а также прошедших профессиональное обучение на производстве, в соответствии с трудовым законодательством и иными нормативными правовыми актами, содержащими нормы трудового права, коллективным договором, соглашениями, ЛНА, трудовым договором могут устанавливаться пониженные нормы выработки.
2. В ч.2 комментируемой статьи подчеркивается невозможность включения в коллективные договоры и соглашения, в трудовой договор условий, ухудшающих правовое положение работников по сравнению с законами и иными нормативными правовыми актами о труде.
В данной норме реализована защитная функция трудового права, выраженная в централизованном установлении стандартов труда, гарантий трудовых прав работников, которые никаким соглашением о труде не могут быть снижены.
Ярким примером реализации комментируемой нормы является ч.2 ст. 232 ТК РФ, которой предусмотрено, что трудовым договором или заключаемыми в письменной форме соглашениями, прилагаемыми к нему, может конкретизироваться материальная ответственность сторон этого договора. При этом договорная ответственность работодателя перед работником не может быть ниже, а работника перед работодателем — выше, чем это предусмотрено ТК РФ или иными федеральными законами.
Если условия, снижающие уровень гарантий прав работников, включены в коллективный договор, соглашение или трудовой договор, то они не подлежат применению.
Иллюстрацией к этому является кассационное определение Верховного суда Удмуртской Республики от 1 сентября 2010 года по делу N 33-2849. В нем установлено, что условие договоров о производственном обучении, в которых указано о полном возмещении стоимости обучения, а не пропорционально фактически не отработанному после окончания обучения времени, ухудшает положение работника по сравнению с положениями ст. 249 ТК РФ, что нарушает ч.2 ст. 232 ТК РФ. Потому данное условие договора признается недействительным.
249 ТК РФ, что нарушает ч.2 ст. 232 ТК РФ. Потому данное условие договора признается недействительным.
Другой комментарий к ст. 9 ТК РФ
1. Комментируемая статья определяет содержание договорно-правового блока источников трудового права. Его образуют: а) коллективные договоры и соглашения, заключаемые между работодателями и профсоюзами либо иными представителями работников; б) индивидуальные трудовые договоры, заключаемые непосредственно работодателями и работниками. Они призваны в условиях рыночной экономики выполнять роль важнейших источников трудового права и нести основную регулирующую нагрузку. С этой целью коллективные договоры и соглашения наряду с обязательственными положениями содержат нормативные положения, которые позволяют им заполнять нишу в системе источников трудового права, оставленную для них нормативными правовыми актами публичных властей.
Эти правовые акты также: а) конкретизируют нормативные источники норм трудового права; б) облекают в надлежащую форму социально-партнерские отношения работодателей и работников; в) служат источником коллективных норм трудового права, адресованных работникам как стороне коллективного договора или соглашения; г) гармонизируют социальные интересы работников и частные интересы работодателя; д) устанавливают пределы локальному нормотворчеству работодателя.
2. Договорно-правовой блок источников трудового права немыслим без включения в него трудового договора, являющегося основным инструментом правового регулирования индивидуальных трудовых отношений в условиях рыночной экономики. Функция индивидуального правового регулирования трудовых отношений выполнялась трудовым договором всегда, однако ее объем в разное время был различен.
В условиях государственно-плановой системы управления производством и трудом основную регулирующую нагрузку несли нормативные правовые акты органов государственного управления. Однако с переходом на рыночные основы хозяйствования государство сложило с себя функции субъекта прямого управления хозяйственной деятельностью и трудом, и отечественная экономика стала функционировать во многом на началах предпринимательской самостоятельности хозяйствующих субъектов и свободного труда работников, используемого на условиях, устанавливаемых самими договаривающимися сторонами при заключении трудовых договоров.
Необходимо подчеркнуть, что индивидуальное регулирование трудовых отношений, осуществляемое с помощью этих договоров, не является разновидностью публичного нормативного правового регулирования, так как в его основе лежит не общая норма права, распространяющаяся на неопределенный круг работников, а персонально определенная договоренность, касающаяся взаимоотношений конкретного работодателя и конкретного работника. Тем не менее трудовой договор сегодня можно и нужно рассматривать как индивидуальный правовой акт, который порождает индивидуальные права и обязанности его сторон и тем самым регулирует их взаимоотношения. В совокупности с локальными нормативными актами и коллективными договорами (соглашениями) трудовые договоры создают у конкретного работодателя определенный правовой режим локального регулирования социально-трудовых отношений. Учитывая эти реалии, закон совершенно правильно включил трудовой договор в число источников договорного регулирования социально-трудовых отношений, а тем самым и в состав элементов системы источников правового регулирования социально-трудовых отношений.
3. Признав договоры и соглашения источниками договорно-правового регулирования социально-трудовых отношений, ТК установил и предел этому виду регулирования посредством законодательной фиксации уровня прав и гарантий работников, который не вправе снижать коллективные договоры, соглашения и трудовые договоры. В случае нарушения этого правила соответствующие условия конкретных договоров или соглашений признаются юридически ничтожными и не подлежат применению.
Консультации и комментарии юристов по ст 9 ТК РФ
Если у вас остались вопросы по статье 9 ТК РФ и вы хотите быть уверены в актуальности представленной информации, вы можете проконсультироваться у юристов нашего сайта.
Задать вопрос можно по телефону или на сайте. Первичные консультации проводятся бесплатно с 9:00 до 21:00 ежедневно по Московскому времени. Вопросы, полученные с 21:00 до 9:00, будут обработаны на следующий день.
1. | Финансирование непредусмотренных расходов | |||
1.2.2.1 | Финансирование расходов, не предусмотренных законом (решением) о бюджете и (или) бюджетной росписью | Ст. 83 БК РФ; ст. 7 Закона Ульяновской области «О бюджетном процессе в Ульяновской области» | Ст. 283, 296 БК РФ;ст. 7 Закона Ульяновской области «О бюджетном процессе в Ульяновской области» | Дополнительные расходы средств бюджетной системы |
1.2.2.2 | Финансирование расходов сверх утвержденных лимитов бюджетных обязательств
| Ст. 226 БК РФ | Ст. 283, 297 БК РФ; ст. 7 Закона Ульяновской области «О бюджетном процессе в Ульяновской области» | Избыточные расходы средств бюджетной системы |
1. | Неправомерное финансирование муниципальных органов власти и муниципальных организаций за счет средств бюджета субъекта РФ | ст. 85, 86 БК РФ |
| Дополнительные расходы средств бюджетной системы Нарушение принципа самостоятельности бюджетов бюджетной системы |
1.2.2.4 | Неправомерное финансирование федеральных органов государственной власти и государственных организаций за счет средств бюджета субъекта РФ или местных бюджетов | Ст. 124 Конституции РФ; ст. 84, 85, 86 БК РФ; положения о соответствующих органах власти |
| Дополнительные расходы средств бюджетной системы Нарушение принципа самостоятельности бюджетов бюджетной системы |
1.2.3 | Иные нарушения при предоставлении бюджетных средств | |||
1. | Финансирование капитальных вложений при отсутствии утвержденной проектно-сметной документации на объекты
| Ст. 80 БК РФ; Ст. 48 Градостроительного кодекса РФ; ст. 14 ФЗ «Об инвестиционной деятельности в РФ, осуществляемой в форме капитальных вложений» |
| Дополнительные расходы средств бюджетной системы [7] |
1.2.3.2 | Неполное, несвоевременное выделение ассигнований на долевое финансирование мероприятий из бюджетов различного уровня бюджетной системы | Закон (решение) о бюджете на соответствующий финансовый год | Ст. 283 БК РФ | Дополнительные расходы средств бюджетной системы |
1.2.3.3 | Превышение полномочий главными распорядителями, распорядителями средств при заключении договоров и соглашений на предоставление бюджетных средств на возвратной основе, государственных или муниципальных гарантий, бюджетных инвестиций | Ст. | Ст. 160, 283 БК РФ; ст. 286 УК РФ | Неэффективное использование средств бюджетной системы |
1.2.3.4 | Неправомерное использование доходов, полученных сверх утвержденных законом о бюджете | Ст. 232, 260 БК РФ; закон (решение) о бюджете на соответствующий финансовый год |
|
|
1.2.4 | Недостатки (неэффективная деятельность) при предоставлении бюджетных средств | |||
1.2.4.1 | Отсутствие контроля со стороны распорядителя бюджетных средств за эффективным и экономным расходованием бюджетных средств подведомственными учреждениями | Ст. 34, ст. | Ст. 160 БК РФ | Неэффективное, неэкономное использование средств бюджетной системы |
1.2.4.2 | Перечисление средств для осуществления регулярных платежей без учета графика таких платежей | Ст. 34 БК РФ | Ст. 160 БК РФ | Неэффективное использование средств бюджетной системы |
1.2.4.3 | Перечисление бюджетных средств на финансирование расходов в сумме большей, чем требуется в действительности (при установлении в ходе контрольных мероприятий, что расчетная потребность в средствах бюджета оказалась завышенной) с учетом возвращенных средств в установленном порядке | Ст. 34, 163 БК РФ | Ст. 160, 283 БК РФ | Неэффективное использование средств бюджетной системы |
1. | Нарушения и недостатки при расходовании бюджетных средств | |||
1.3.1 | Нецелевое использование средств бюджета | |||
1.3.1.1 | Использование бюджетных средств на цели, не предусмотренные законом (решением) о бюджете, бюджетной росписью и уведомлениями о бюджетных ассигнованиях | Ст. 38, 70, 161, 163 БК РФ | Ст.160, 231, 283, 289 БК РФ; ст. 15.14 КоАП РФ; ст. 285.1 УК РФ; ст. 1 Закона Ульяновской области «Об административной ответственности за нарушения бюджетного законодательства в Ульяновской области» | Неэффективное использование средств бюджетной системы Нарушение принципа адресности и целевого характера бюджетных средств бюджетной системы |
1. | Использование бюджетных средств на оплату расходов, не предусмотренных сметой доходов и расходов | Ст. 38, 70, 161, 163 БК РФ | Ст.160, 231, 283, 289 БК РФ; ст. 15.14 КоАП РФ; ст. 285.1 УК РФ; ст.1 Закона Ульяновской области «Об административной ответственности за нарушения бюджетного законодательства в Ульяновской области» | Неэффективное использование средств бюджетной системы Нарушение принципа адресности и целевого характера бюджетных средств бюджетной системы |
1.3.1.3 | Расходование бюджетных средств не по коду экономической (функциональной) классификации расходов бюджетов РФ, по которому было произведено финансирование (несанкционированное перемещение бюджетных средств по кодам экономической (функциональной) классификации расходов бюджетов РФ) | Ст. [8] | Ст. 160, 283, 289 БК РФ; ст. 15.14 КоАП РФ; ст.285.1 УК РФ ст. 1 Закона Ульяновской области «Об административной ответственности за нарушения бюджетного законодательства в Ульяновской области» | Нарушение регламентированного режима использования средств бюджетной системы |
1.3.1.4 | Использование субвенций, субсидий и иных средств (за исключением бюджетных кредитов/ссуд) не в соответствии с условиями их предоставления
| Ст. 6, 38, 78 БК РФ; НПА Ульяновской области | Ст. 78, 231, 283, 289 БК РФ; ст. 15.14 КоАП РФ; ст. 1 Закона Ульяновской области «Об административной ответственности за нарушения бюджетного законодательства в Ульяновской области» | Неэффективное использование средств бюджетной системы Нарушение принципа адресности и целевого характера бюджетных средств бюджетной системы |
1. | Иное неправомерное использование бюджетных средств | |||
1.3.2.1 | Использование бюджетных средств на финансирование затрат, не предусмотренных проектно-сметной документацией, необоснованное завышение объемов работ | Ст. 38, 163 БК РФ; закон (решение) о бюджете на соответствующий финансовый год | Ст. 160, 283, 289 БК РФ; ст. 15.14 КоАП РФ; ст.285.1 УК РФ | Избыточные расходы средств бюджетной системы |
1.3.2.2 | Использование бюджетных средств на оплату расходов, которые должны осуществляться за счет средств, поступающих из бюджетов других уровней, за исключением случаев, когда федеральным законодательством предусмотрен порядок финансирования федеральных полномочий через возмещение фактически осуществленных затрат | Ст. | Ст.160, 231, 283, 289 БК РФ; ст. 15.14 КоАП РФ; ст. 285.1 УК РФ | Дополнительные расходы средств бюджетной системы Нарушение принципа самостоятельности бюджетов |
1.3.2.3 | Использование бюджетных средств на оплату расходов, которые должны осуществляться за счет внебюджетных источников | Ст. 38, 161, 163 БК РФ | Ст.160, 231, 283, 289 БК РФ; ст. 15.14 КоАП РФ; ст. 285.1 УК РФ | Дополнительные расходы средств бюджетной системы |
1.3.2.4 | Использование бюджетных средств на оплату штрафов и иных санкций, наложенных на должностных лиц | Ст. 38, 161, 163 БК РФ | Ст.160, 231, 283, 289 БК РФ; ст. 15.14 КоАП РФ | Дополнительные расходы средств бюджетной системы |
1. | Выплаты из бюджета пособий, компенсаций, субсидий и других денежных выплат лицам, не имеющим правовых оснований для их получения | Ст. 38, 70, 75, 161, 163 БК РФ; НПА Ульяновской области, муниципальные правовые акты | Ст. 78, 160, 231, 283, 289 БК РФ; ст. 15.14 КоАП РФ; ст. 285.1 УК РФ | Дополнительные расходы средств бюджетной системы Необоснованное расходование средств бюджетной системы |
1.3.2.6 | Использование бюджетных средств на оплату работ и услуг, не связанных с деятельностью бюджетного учреждения | Ст. 38, 70, 161, 163 БК РФ | Ст. 160, 231, 283, 289 БК РФ; ст. 15.14 КоАП РФ; ст. 285.1 УК РФ | Дополнительные расходы средств бюджетной системы |
1.3.2.7 | Использование бюджетных средств на оплату коммунальных услуг, оказанных арендаторам, без соответствующего возмещения арендаторами | Ст. | Ст. 160, 231, 283, 289 БК РФ; ст. 15.14 КоАП РФ; ст. 285.1 УК РФ | Дополнительные расходы средств бюджетной системы |
1.3.2.8 | Расходование бюджетных средств без оправдательных документов | Ст. 38 БК РФ; ст. 9 ФЗ «О бухгалтерском учете» | Ст. 160, 231, 283, 289 БК РФ; ст. 18 ФЗ «О бухгалтерском учете»; ст. 15.14, 19.23 КоАП РФ | Необоснованное расходование средств бюджетной системы |
1.3.2.9 | Расходование бюджетных средств по документам, не соответствующим действительности (поддельным) | ст. 9 ФЗ «О бухгалтерском учете» | ст. 18 ФЗ «О бухгалтерском учете»; ст. 292, 327 УК РФ | Утрата средств бюджетной системы |
1. | Неправомерное принятие и подтверждение обязательств, расходование сверх утвержденных показателей | |||
1.3.3.1 | Неправомерное принятие денежных обязательств (сверх доведенных лимитов бюджетных обязательств, по истечении установленного срока) | Ст. 225, 242, 250 БК РФ | 297 БК РФ | Избыточные расходы средств бюджетной системы |
1.3.3.2 | Подтверждение денежных обязательств, не подлежащих принятию | Ст. 226, 242 БК РФ | Ст. 226 БК РФ | Дополнительные расходы средств бюджетной системы |
1.3.3.3 | Расходование бюджетных средств сверх показателей, утвержденных законом (решением) о бюджете, сводной бюджетной росписью, сметой доходов и расходов, сверх доведенных лимитов бюджетных обязательств | Ст. | 297 БК РФ | Избыточные расходы средств бюджетной системы |
1.3.3.4 | Переплаты и непредусмотренные законодательством выплаты заработной платы (денежного содержания, довольствия и пр.)
| Ст. 38, 70 БК РФ; ст.57, 91, 96, 103, 104 ТК РФ Ст. 50 ФЗ «О государственной гражданской службе РФ»; Ст. 16 Закона Ульяновской области «О государственных должностях Ульяновской области»; Ст. 9 Закона Ульяновской области «О государственной гражданской службе Ульяновской области»; Ст. 16 ФЗ «Об основах муниципальной службы в РФ»; Закон Ульяновской области «Об установлении предельных нормативов оплаты труда лиц, замещающих отдельные муниципальные должности муниципальных образований Ульяновской области»; другие НПА Ульяновской области, муниципальные правовые акты | ст. | Избыточные расходы средств бюджетной системы |
— необоснованное завышение должностных окладов (завышение процентных надбавок за выслугу лет, за классный чин) | ||||
— установление техническим служащим окладов и размера ежемесячного денежного поощрения в размере, предусмотренном для государственных гражданских служащих | ||||
— нарушения в штатной численности учреждения (искусственное завышение численности, наличие большого количества вакантных ставок, что позволяет устанавливать работникам надбавки за совмещение должностей и разовые премии) | ||||
1.3.3.5 | Сверхнормативные расходы
| Ст. 38, 70, 161, 163 БК РФ | Ст. 160, 283, 289, 297 БК РФ; ст. 15.14 КоАП РФ; ст. | Избыточные расходы средств бюджетной системы |
— превышение лимитов на поездки автотранспорта | ||||
| — нарушения в потреблении и оплате электроэнергии за счет несанкционированного подключения сторонних организаций; без надлежащего оформления соответствующих документов и средств измерения; |
|
|
|
— нарушения в потреблении и оплате водоснабжения – питание питьевой водой сторонних организаций без надлежащего оформления соответствующих документов и средств измерения; | ||||
— завышение сметы расходов на ГСМ в сравнении с фактическим потреблением | ||||
— оплата ГСМ для автомобилей не принадлежащих учреждениям | ||||
— отсутствие в учреждениях санкций за превышение лимитов на разговоры по сотовым телефонам и за личные междугородние переговоры; | ||||
— оплата услуг сотовой связи за телефонные номера не принадлежащие учреждению | ||||
1. | Неправомерное неподтверждение обязательств, неплатежи по бюджетным обязательствам | |||
1.3.4.1 | Неправомерный отказ в подтверждении бюджетных обязательств получателям бюджетных средств | Ст. 226 БК РФ | Ст. 226, 283, 306 БК РФ Ст. 19.7 КоАП | Дезорганизация бюджетного процесса |
1.3.4.2 | Несвоевременное подтверждение бюджетных обязательств | Ст. 226 БК РФ | Ст. 226, 283, 306 БК РФ | Дезорганизация бюджетного процесса |
1.3.4.3 | Неплатежи, несвоевременные платежи по бюджетным обязательствам, влекущие выплату пени, штрафов | Ст. | Ст. 283, 306 БК РФ; ст. 395 ГК РФ; ст. 142, 236 ТК РФ; ст. 5.27, 5.31 КоАП РФ | Дополнительные расходы средств бюджетной системы |
1.3.4.4 | Незачисление бюджетных средств на счета получателей бюджетных средств при условии достаточности средств на едином счете бюджета (несвоевременное отражение на лицевых счетах) | Ст. 163 БК РФ | Ст. 283, 293, 305 БК РФ | Дополнительные расходы средств бюджетной системы [9] |
1.3.4.5 | Несвоевременное осуществление платежей по подтвержденным бюджетным обязательствам при условии достаточности средств на едином счете бюджета | Ст. 163 БК РФ | Ст. 283, 306 БК РФ | Дополнительные расходы средств бюджетной системы [10] |
1. | Необеспечение сохранности, возврата бюджетных средств | |||
1.3.5.1 | Невостребование с работника бюджетного учреждения – подотчетного лица задолженности по бюджетным средствам, выданным ему под отчет | Ст. 137 ТК РФ; п.157-160, п.17 приложения № 3 Инструкции по бюджетному учету №70н [11] — до 01.01.2006 и п.155 Инструкции по бюджетному учету № 25н [12] — с 01.01.2006; п. 11 Порядка ведения кассовых операций в РФ [13] | Ст. 18 ФЗ «О бухгалтерском учете» | Недопоступление средств в бюджетную систему |
1.3.5.2 | Недостача государственных (муниципальных) денежных средств | Ст. 7 ФЗ «О бухгалтерском учете»; п. | Ст. 243 ТК РФ | Утрата средств бюджетной системы |
 2.2
2.2 2.2.3
2.2.3 2.3.1
2.3.1 158, 159 БК РФ
158, 159 БК РФ 159 БК РФ
159 БК РФ 3
3 3.1.2
3.1.2 38, 161, 163 БК РФ; ФЗ «О бюджетной классификации РФ»; Указания о порядке применения бюджетной классификации РФ
38, 161, 163 БК РФ; ФЗ «О бюджетной классификации РФ»; Указания о порядке применения бюджетной классификации РФ 3.2
3.2 31, 84, 85, 86 БК РФ
31, 84, 85, 86 БК РФ 3.2.5
3.2.5 38, 70, 161, 163 БК РФ
38, 70, 161, 163 БК РФ 3.3
3.3 225, 226 БК РФ; закон (решение) о бюджете на соответствующий финансовый год
225, 226 БК РФ; закон (решение) о бюджете на соответствующий финансовый год 160, 283, 289 БК РФ; ст. 15.14 КоАП РФ; ст.285.1 УК РФ
160, 283, 289 БК РФ; ст. 15.14 КоАП РФ; ст.285.1 УК РФ 285.1 УК РФ
285.1 УК РФ 3.4
3.4 136 ТК РФ
136 ТК РФ 3.5
3.5 п. 161-164 Инструкции по бюджетному учету №70н – до 01.01.2006; Инструкция по бюджетному учету №25н – с 01.01.2006
п. 161-164 Инструкции по бюджетному учету №70н – до 01.01.2006; Инструкция по бюджетному учету №25н – с 01.01.2006официальные выходные и праздничные дни в России
https://ria.ru/20220201/prazdniki-1770508979.html
Как отдыхаем в марте 2022 года: перенос выходных и сокращенные дни
Как отдыхаем в марте 2022: официальные выходные и праздничные дни в России
Как отдыхаем в марте 2022 года: перенос выходных и сокращенные дни
Международный женский день 8 марта в 2022 году выпадает на вторник. Как отдыхаем в марте, сколько будет официальных нерабочих дней в месяце и какие будут… РИА Новости, 01.02.2022
2022-02-01T16:25
2022-02-01T16:25
2022-02-01T18:09
россия
общество
праздники
8 марта
празднование 8 марта
/html/head/meta[@name=’og:title’]/@content
/html/head/meta[@name=’og:description’]/@content
https://cdnn21. img.ria.ru/images/07e6/02/01/1770501123_0:0:3072:1728_1920x0_80_0_0_2315469d93b05fabe405dc3dc571f0ba.jpg
img.ria.ru/images/07e6/02/01/1770501123_0:0:3072:1728_1920x0_80_0_0_2315469d93b05fabe405dc3dc571f0ba.jpg
МОСКВА, 1 фев — РИА Новости. Международный женский день 8 марта в 2022 году выпадает на вторник. Как отдыхаем в марте, сколько будет официальных нерабочих дней в месяце и какие будут переносы выходных в связи с праздником — в материале РИА Новости.Как отдыхаем в марте 2022Выходные и праздничные дни в России определены статьей 112 Трудового кодекса. Международный женский день, который ежегодно отмечается в марте, по закону нерабочий. При этом власти устанавливают, сколько будет длиться отдых, приуроченный к 8 марта, и переносятся ли выходные.Выходные дниНа первый весенний месяц 2022 года приходится девять праздничных и выходных дней, в том числе один государственный праздник — Международный женский день.Выходные дни в марте 2022:Праздник 8 мартаВ России Международный женский день впервые отметили в 1913 году. Тогда он был посвящен солидарности женщин в борьбе за равные права. В Советском Союзе день 8 марта так или иначе праздновался всегда, однако лишь с 1966 года, по указу Президиума Верховного Совета СССР от 8 мая 1965 года, этот день стал официально праздничным и нерабочим. После распада Советского Союза день 8 марта остался в перечне государственных праздников Российской Федерации, согласно статье 112 Трудового кодекса РФ. Сейчас эта дата признана нерабочим праздничным днем во всех странах бывшего СССР, кроме Латвии, Литвы и Эстонии.Переносы выходных днейПоскольку 8 марта в этом году приходится на вторник, правительство приняло решение перенести выходной с субботы, 5 марта, на понедельник, 7 марта. Соответственно, россиян ждут трехдневные выходные дни с 6 по 8 марта (с субботы по вторник).Производственный календарьПроизводственный календарь определяет количество рабочих, праздничных и выходных дней. Помимо этого, в нем указана норма рабочего времени.Осенью 2021 года правительство России утвердило график на 2022 год, согласно которому 8 марта 2022 – нерабочий день.Сколько рабочих днейНа основании статьи 95 Трудового кодекса Российской Федерации длительность рабочего дня, предшествующего нерабочему праздничному дню, уменьшается на один час. Но нередки случаи, когда работодатель устанавливает другой распорядок в организации.
После распада Советского Союза день 8 марта остался в перечне государственных праздников Российской Федерации, согласно статье 112 Трудового кодекса РФ. Сейчас эта дата признана нерабочим праздничным днем во всех странах бывшего СССР, кроме Латвии, Литвы и Эстонии.Переносы выходных днейПоскольку 8 марта в этом году приходится на вторник, правительство приняло решение перенести выходной с субботы, 5 марта, на понедельник, 7 марта. Соответственно, россиян ждут трехдневные выходные дни с 6 по 8 марта (с субботы по вторник).Производственный календарьПроизводственный календарь определяет количество рабочих, праздничных и выходных дней. Помимо этого, в нем указана норма рабочего времени.Осенью 2021 года правительство России утвердило график на 2022 год, согласно которому 8 марта 2022 – нерабочий день.Сколько рабочих днейНа основании статьи 95 Трудового кодекса Российской Федерации длительность рабочего дня, предшествующего нерабочему праздничному дню, уменьшается на один час. Но нередки случаи, когда работодатель устанавливает другой распорядок в организации. В марте 2022 года 22 рабочих дня, включая один сокращенный (5 марта).Норма рабочего времени в мартеНормы продолжительности рабочего времени в марте:Могут ли заставить работать в праздничные выходныеСогласно статье 113 Трудового кодекса Российской Федерации работа в праздничные и выходные дни запрещена. Поэтому работодатель не вправе заставить сотрудника выйти на работу с 6 по 8 марта. Исключением являются случаи, когда сотрудник согласился сделать это по просьбе руководителя. В этом случае работнику необходимо подписать письменное согласие, а начальник должен издать соответствующий приказ и ознакомить с ним персонал.Когда выгодно взять отпуск в мартеЧтобы уйти в отпуск «выгодно» с точки зрения финансов, нужно иметь в виду, что согласно ТК РФ праздничные дни не оплачиваются и в дни отпуска не включаются.Соответственно, если брать отпуск в тот месяц, в котором много выходных и праздничных дней, компенсация будут меньше.Март считается не самым финансово выгодным месяцем для отпуска. Но стоит учитывать, что ежегодный отдых, запланированный на начало марта, можно продлить на три дня.
В марте 2022 года 22 рабочих дня, включая один сокращенный (5 марта).Норма рабочего времени в мартеНормы продолжительности рабочего времени в марте:Могут ли заставить работать в праздничные выходныеСогласно статье 113 Трудового кодекса Российской Федерации работа в праздничные и выходные дни запрещена. Поэтому работодатель не вправе заставить сотрудника выйти на работу с 6 по 8 марта. Исключением являются случаи, когда сотрудник согласился сделать это по просьбе руководителя. В этом случае работнику необходимо подписать письменное согласие, а начальник должен издать соответствующий приказ и ознакомить с ним персонал.Когда выгодно взять отпуск в мартеЧтобы уйти в отпуск «выгодно» с точки зрения финансов, нужно иметь в виду, что согласно ТК РФ праздничные дни не оплачиваются и в дни отпуска не включаются.Соответственно, если брать отпуск в тот месяц, в котором много выходных и праздничных дней, компенсация будут меньше.Март считается не самым финансово выгодным месяцем для отпуска. Но стоит учитывать, что ежегодный отдых, запланированный на начало марта, можно продлить на три дня. Работодатель не вправе отказать сотруднику в этом.
Работодатель не вправе отказать сотруднику в этом.
https://ria.ru/20210308/prazdnik-1600047359.html
россия
РИА Новости
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
2022
РИА Новости
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
Новости
ru-RU
https://ria.ru/docs/about/copyright.html
https://xn--c1acbl2abdlkab1og.xn--p1ai/
РИА Новости
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
https://cdnn21.img.ria.ru/images/07e6/02/01/1770501123_151:0:2882:2048_1920x0_80_0_0_d92f10b969f119763241ca5024cdf075.jpgРИА Новости
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og. xn--p1ai/awards/
xn--p1ai/awards/
РИА Новости
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
россия, общество, праздники, 8 марта, празднование 8 марта
МОСКВА, 1 фев — РИА Новости. Международный женский день 8 марта в 2022 году выпадает на вторник. Как отдыхаем в марте, сколько будет официальных нерабочих дней в месяце и какие будут переносы выходных в связи с праздником — в материале РИА Новости.
Как отдыхаем в марте 2022
Выходные и праздничные дни в России определены статьей 112 Трудового кодекса. Международный женский день, который ежегодно отмечается в марте, по закону нерабочий. При этом власти устанавливают, сколько будет длиться отдых, приуроченный к 8 марта, и переносятся ли выходные.Выходные дни
На первый весенний месяц 2022 года приходится девять праздничных и выходных дней, в том числе один государственный праздник — Международный женский день.
Выходные дни в марте 2022:
—
6- 8 марта;—
12-13 марта;—
19-20 марта;—
26-27 марта.
Праздник 8 марта
В России Международный женский день впервые отметили в 1913 году. Тогда он был посвящен солидарности женщин в борьбе за равные права. В Советском Союзе день 8 марта так или иначе праздновался всегда, однако лишь с 1966 года, по указу Президиума Верховного Совета СССР от 8 мая 1965 года, этот день стал официально праздничным и нерабочим.8 марта 2021, 04:03
Международный женский деньПосле распада Советского Союза день 8 марта остался в перечне государственных праздников Российской Федерации, согласно статье 112 Трудового кодекса РФ. Сейчас эта дата признана нерабочим праздничным днем во всех странах бывшего СССР, кроме Латвии, Литвы и Эстонии.Переносы выходных дней
Поскольку 8 марта в этом году приходится на вторник, правительство приняло решение перенести выходной с субботы, 5 марта, на понедельник, 7 марта. Соответственно, россиян ждут трехдневные выходные дни с 6 по 8 марта (с субботы по вторник).
Соответственно, россиян ждут трехдневные выходные дни с 6 по 8 марта (с субботы по вторник).
Производственный календарь
Производственный календарь определяет количество рабочих, праздничных и выходных дней. Помимо этого, в нем указана норма рабочего времени.
Осенью 2021 года правительство России утвердило график на 2022 год, согласно которому 8 марта 2022 – нерабочий день.
Сколько рабочих дней
На основании статьи 95 Трудового кодекса Российской Федерации длительность рабочего дня, предшествующего нерабочему праздничному дню, уменьшается на один час. Но нередки случаи, когда работодатель устанавливает другой распорядок в организации.В марте 2022 года 22 рабочих дня, включая один сокращенный (5 марта).
Норма рабочего времени в марте
Нормы продолжительности рабочего времени в марте:
—
при 40-часовой рабочей неделе – 175 часов;—
при 36-часовой рабочей неделе – 157,4 часа;—
при 24-часовой рабочей неделе – 104,6 часа.

Могут ли заставить работать в праздничные выходные
Согласно статье 113 Трудового кодекса Российской Федерации работа в праздничные и выходные дни запрещена. Поэтому работодатель не вправе заставить сотрудника выйти на работу с 6 по 8 марта. Исключением являются случаи, когда сотрудник согласился сделать это по просьбе руководителя. В этом случае работнику необходимо подписать письменное согласие, а начальник должен издать соответствующий приказ и ознакомить с ним персонал.Когда выгодно взять отпуск в марте
Чтобы уйти в отпуск «выгодно» с точки зрения финансов, нужно иметь в виду, что согласно ТК РФ праздничные дни не оплачиваются и в дни отпуска не включаются.Соответственно, если брать отпуск в тот месяц, в котором много выходных и праздничных дней, компенсация будут меньше.
Март считается не самым финансово выгодным месяцем для отпуска. Но стоит учитывать, что ежегодный отдых, запланированный на начало марта, можно продлить на три дня. Работодатель не вправе отказать сотруднику в этом.
Смоделированное радиационное воздействие прямого аэрозольного воздействия с оценкой нескольких наблюдений
19 февраля 2009 г.
19 февраля 2009 г.
Г. Мире 1,2 ,Т. Ф. Берглен 2,3 ,М. Джонсруд 3 ,К. Р. Хойл 2 , Т. К. Бернтсен 1,2 ,С. А. Кристофер 4 , Д. В. Фэйи 5,6 , И.С. А. Исаксен 1,2 ,Т. А. Джонс 4 , Р. А. Кан 7 , Н. Леб 8 , стр. Куинн 9 ,Л. Ремер 10 ,Дж. П. Шварц 5,6 и К. Э. Иттри 3 Г. Мире и соавт. Г. Мире 1,2 ,Т. Ф. Берглен 2,3 ,М. Джонсруд 3 ,К. Р. Хойл 2 , Т. К. Бернтсен 1,2 ,С. А. Кристофер 4 , Д. В. Фэйи 5,6 , И. С. А. Исаксен 1,2 ,Т.А. Джонс 4 , Р. А. Кан 7 , Н. Леб 8 , стр. Куинн 9 ,Л. Ремер 10 ,Дж. П. Шварц 5,6 и К. Э. Иттри 3
В. Фэйи 5,6 , И. С. А. Исаксен 1,2 ,Т.А. Джонс 4 , Р. А. Кан 7 , Н. Леб 8 , стр. Куинн 9 ,Л. Ремер 10 ,Дж. П. Шварц 5,6 и К. Э. Иттри 3 - 1 Центр международных исследований климата и окружающей среды в Осло, Осло, Норвегия
- 2 Департамент наук о Земле, Университет Осло, Осло, Норвегия
- 3 Норвежский институт исследований воздуха (NILU), Кьеллер, Норвегия
- 4 Департамент атмосферных наук Университета Алабамы в Хантсвилле, Хантсвилл, Алабама, США
- 5 Отдел химических наук, Лаборатория исследования системы Земли, NOAA, Боулдер, Колорадо, США
- 6 Кооператив Институт исследований в области наук об окружающей среде, Университет Колорадо, Боулдер, Колорадо, США
- 7 Лаборатория атмосфер, Центр космических полетов имени Годдарда НАСА, Гринбелт, Мэриленд, США
- 8 Центр атмосферных исследований НАСА в Лэнгли (LaRC), Хэмптон, Вирджиния, США
- 9 NOAA PMEL, Сиэтл, США
- 10 Лаборатория атмосфер, Центр космических полетов имени Годдарда НАСА (GSFC), Гринбелт, Мэриленд, США
- 1 Центр международных исследований климата и окружающей среды в Осло, Осло, Норвегия
- 2 Департамент наук о Земле, Университет Осло, Осло, Норвегия
- 3 Норвежский институт исследований воздуха (NILU), Кьеллер, Норвегия
- 4 Департамент атмосферных исследований Университета Алабамы в Хантсвилле, Хантсвилл, Алабама, США
- 5 Отдел химических наук, Лаборатория исследования системы Земли, NOAA, Боулдер, Колорадо, США
- 6 Институт исследований в области наук об окружающей среде, Университет Колорадо, Боулдер, Колорадо, США
- 7 Лаборатория атмосфер, Центр космических полетов имени Годдарда НАСА, Гринбелт, Мэриленд, США
- 8 NASA Langley Atmospheric Research h Center (LaRC), Хэмптон, Вирджиния, США
- 9 NOAA PMEL, Сиэтл, США
- 10 Лаборатория атмосфер, Центр космических полетов имени Годдарда НАСА (GSFC), Гринбелт, Мэриленд, США
 – Начало обсуждения: 7 июля 2008 г. – Опубликовано: 19 февраля 2009 г.
– Начало обсуждения: 7 июля 2008 г. – Опубликовано: 19 февраля 2009 г. Модель глобального аэрозоля высокого разрешения (Oslo CTM2), основанная на метеорологических данных и позволяющая проводить сравнение с различными наблюдениями за аэрозолями, используется для имитации радиационного воздействия (РВ) прямого аэрозольного воздействия.Модель имитирует все основные компоненты аэрозоля, включая несколько вторичных компонентов, таких как нитраты и вторичный органический углерод. Модель воспроизводит основные особенности химического состава и размеров, наблюдаемые во время крупных аэрозольных кампаний. Хотя химический состав смоделированного сульфата лучше всего сравнивается с наземными измерениями над сушей, для других соединений систематических различий обнаружено не было. Смоделированная оптическая толщина аэрозоля (AOD) сравнивается с данными дистанционного зондирования, полученными наземными данными AERONET, а также данными спутников MODIS и MISR.Чтобы обрести уверенность в моделировании аэрозолей, мы проверили его способность воспроизводить ежедневную изменчивость содержания аэрозолей, и это хорошо работает во многих регионах; однако мы также определили некоторые места, где необходимы улучшения модели. Среднегодовая региональная картина АОТ по аэрозольной модели в целом аналогична данным AERONET и спутниковым поискам (в основном в пределах 10–20%). Мы заметили значительное улучшение по сравнению с коллекцией 4 MODIS по сравнению с коллекцией 5 по сравнению с данными AERONET.Спутниковые оценки радиационного воздействия аэрозолей над океаном в условиях ясного неба значительно различаются в региональном масштабе (почти в два раза), а также в среднем глобальном масштабе. Oslo CTM2 имеет аэрозольный радиационный эффект, близкий к среднему значению оценок, полученных со спутника. Мы получаем радиационное воздействие (RF) прямого аэрозольного воздействия, равное −0,35 Вт·м −2 в нашем базовом случае. Реализация простого подхода для учета внутренней смеси черного углерода (ЧУ) приводит к общему RF, равному −0.28 Wm −2 . Наши результаты подчеркивают важность углеродистых частиц, производящих более сильное индивидуальное радиочастотное излучение, чем предполагалось в недавней оценке МГЭИК; однако чистый RF менее отличается.
Среднегодовая региональная картина АОТ по аэрозольной модели в целом аналогична данным AERONET и спутниковым поискам (в основном в пределах 10–20%). Мы заметили значительное улучшение по сравнению с коллекцией 4 MODIS по сравнению с коллекцией 5 по сравнению с данными AERONET.Спутниковые оценки радиационного воздействия аэрозолей над океаном в условиях ясного неба значительно различаются в региональном масштабе (почти в два раза), а также в среднем глобальном масштабе. Oslo CTM2 имеет аэрозольный радиационный эффект, близкий к среднему значению оценок, полученных со спутника. Мы получаем радиационное воздействие (RF) прямого аэрозольного воздействия, равное −0,35 Вт·м −2 в нашем базовом случае. Реализация простого подхода для учета внутренней смеси черного углерода (ЧУ) приводит к общему RF, равному −0.28 Wm −2 . Наши результаты подчеркивают важность углеродистых частиц, производящих более сильное индивидуальное радиочастотное излучение, чем предполагалось в недавней оценке МГЭИК; однако чистый RF менее отличается. Оценивается значительное RF от вторичных органических аэрозолей (SOA) (около -0,1 Wm -2 ). SOA также способствует сильному преобладанию вторичных видов аэрозолей в составе аэрозолей над сушей. Сочетание моделирования чувствительности и оценки модели показывает, что RF довольно устойчив и вряд ли будет намного сильнее, чем в нашей наилучшей оценке.
Оценивается значительное RF от вторичных органических аэрозолей (SOA) (около -0,1 Wm -2 ). SOA также способствует сильному преобладанию вторичных видов аэрозолей в составе аэрозолей над сушей. Сочетание моделирования чувствительности и оценки модели показывает, что RF довольно устойчив и вряд ли будет намного сильнее, чем в нашей наилучшей оценке.
Иммунный ответ против вариантов SARS-CoV-2 после гетерологичной и гомологичной вакцинации ChAdOx1 nCoV-19/BNT162b2 — это продолжающееся проспективное обсервационное исследование, в котором отслеживается иммуноглобулин IgG против SARS-CoV-2 и иммунный ответ у 1493 медицинских работников Ганноверской медицинской школы и у лиц, потенциально контактировавших с SARS-CoV-2 (ссылки.
18,22 ). Поправка от декабря 2020 года позволила нам изучить иммунные реакции после вакцинации против COVID-19. В соответствии с немецкими правилами, медицинские работники имели приоритет для вакцинации против SARS-CoV-2, а медицинские работники в Ганноверской медицинской школе получили первые дозы либо вакцины BNT после 6 января, либо вакцины ChAd после 16 февраля 2021 года. через 21 день после праймирования BNT и через 2–3 месяца после праймирования ChAd. Бустерная вакцинация HCP, примированных ChAd, началась 3 мая 2021 г., и люди могли выбрать получение либо ChAd, либо BNT для повторной вакцинации.Мы предположили, что около 25% всех вакцинированных ChAd предпочтут гомологичную бустерную иммунизацию. Расчет мощности, выполненный с помощью G*Power (v3.1.9.6), показал, что размер выборки из 30 человек в каждой группе достаточен для выявления клинически значимых различий внутри каждой группы, при условии, что IgG, специфичные к шиповидным белкам, удваиваются с момента первой вакцинации. (среднее, 95 отн. ед. (RU) мл -1 , при стандартном отклонении 113 RU мл -1 ) и при использовании двустороннего парного t -критерия различий между средними с 95% мощностью и 1% уровень значимости.На основании приведенных выше расчетов и ожидаемого показателя потери для наблюдения в размере 10% 130 вакцинированных ChAd вакцинированных из когорты исследования CoCo были приглашены сдать кровь до начала бустерной вакцинации в начале мая 2021 года.
через 21 день после праймирования BNT и через 2–3 месяца после праймирования ChAd. Бустерная вакцинация HCP, примированных ChAd, началась 3 мая 2021 г., и люди могли выбрать получение либо ChAd, либо BNT для повторной вакцинации.Мы предположили, что около 25% всех вакцинированных ChAd предпочтут гомологичную бустерную иммунизацию. Расчет мощности, выполненный с помощью G*Power (v3.1.9.6), показал, что размер выборки из 30 человек в каждой группе достаточен для выявления клинически значимых различий внутри каждой группы, при условии, что IgG, специфичные к шиповидным белкам, удваиваются с момента первой вакцинации. (среднее, 95 отн. ед. (RU) мл -1 , при стандартном отклонении 113 RU мл -1 ) и при использовании двустороннего парного t -критерия различий между средними с 95% мощностью и 1% уровень значимости.На основании приведенных выше расчетов и ожидаемого показателя потери для наблюдения в размере 10% 130 вакцинированных ChAd вакцинированных из когорты исследования CoCo были приглашены сдать кровь до начала бустерной вакцинации в начале мая 2021 года. Планирование визитов для вакцинации координировалось независимая бригада по вакцинации в зависимости от наличия вакцины. После письменного информированного согласия мы получили образцы периферической крови путем венепункции. В порядке очереди мы провели формальный статистический анализ после того, как по крайней мере 30 человек в каждой группе получили ревакцинацию и прошли 13-й день после ревакцинации.Один человек с предшествующей инфекцией SARS-CoV-2, что определялось положительным анти-SARS-CoV-2 нуклеокапсидным (NCP) IgG до вакцинации, был исключен из этого анализа. Участниками были 25 % мужчин и 75 % женщин, средний возраст которых составлял 38 лет (от 19 до 64 лет), и, таким образом, они были репрезентативными для всех вакцинированных в исследовании CoCo (72 % женщин, 28 % мужчин; средний возраст 40 лет, диапазон 19–67 лет). После сбора крови мы отделяли плазму от крови с ЭДТА или литий-гепарином (S-Monovette, Sarstedt) и хранили ее при температуре -80 °C до использования.Для стимуляции пулами пептидов SARS-CoV-2 мы использовали мононуклеарные клетки цельной крови или периферической крови (PBMC) из образцов цельной крови, выделенных центрифугированием в градиенте фиколла.
Планирование визитов для вакцинации координировалось независимая бригада по вакцинации в зависимости от наличия вакцины. После письменного информированного согласия мы получили образцы периферической крови путем венепункции. В порядке очереди мы провели формальный статистический анализ после того, как по крайней мере 30 человек в каждой группе получили ревакцинацию и прошли 13-й день после ревакцинации.Один человек с предшествующей инфекцией SARS-CoV-2, что определялось положительным анти-SARS-CoV-2 нуклеокапсидным (NCP) IgG до вакцинации, был исключен из этого анализа. Участниками были 25 % мужчин и 75 % женщин, средний возраст которых составлял 38 лет (от 19 до 64 лет), и, таким образом, они были репрезентативными для всех вакцинированных в исследовании CoCo (72 % женщин, 28 % мужчин; средний возраст 40 лет, диапазон 19–67 лет). После сбора крови мы отделяли плазму от крови с ЭДТА или литий-гепарином (S-Monovette, Sarstedt) и хранили ее при температуре -80 °C до использования.Для стимуляции пулами пептидов SARS-CoV-2 мы использовали мононуклеарные клетки цельной крови или периферической крови (PBMC) из образцов цельной крови, выделенных центрифугированием в градиенте фиколла.
pVNT
pVNT были выполнены в отделении биологии инфекций Немецкого центра приматов в Геттингене, как описано ранее 12 . Вкратце, рабдовирусные псевдотипированные частицы были получены в клетках 293T, трансфицированных для экспрессии желаемого варианта SARS-CoV-2-S, инокулированных VSV*DG-FLuc, вектором вируса везикулярного стоматита (VSV) с дефицитом репликации, который кодирует усиленный зеленый флуоресцентный белок. и люциферазу светлячка (FLuc) вместо белка VSV-G (любезно предоставлено Гертом Циммером, Институт вирусологии и иммунологии, Миттельхойзерн, Швейцария).Полученные псевдочастицы собирали, очищали от клеточного дебриса центрифугированием и хранили при -80°С до использования. Для экспериментов по нейтрализации равные объемы псевдотипированных частиц и инактивированных нагреванием (56 °С, 30 мин) образцов плазмы, серийно разведенных в культуральной среде, смешивали и инкубировали в течение 30 мин при 37 °С. После этого образцы вместе с псевдотипированными частицами, не подвергшимися воздействию плазмы, использовались для экспериментов по трансдукции. Анализ проводили в 96-луночных планшетах, в которые клетки Vero инокулировали соответствующими смесями псевдотипированных частиц/плазмы.Эффективность трансдукции анализировали через 16–18 часов после инокуляции путем измерения активности FLuc в лизированных клетках (Cell Culture Lysis Reagent, Promega) с использованием коммерческого субстрата (Beetle-Juice, PJK) и планшетного люминометра (Hidex Sense Microplate Reader, Hidex). с программным обеспечением для считывания микропланшетов Hidex Sense (v0.5.41.0).
Анализ проводили в 96-луночных планшетах, в которые клетки Vero инокулировали соответствующими смесями псевдотипированных частиц/плазмы.Эффективность трансдукции анализировали через 16–18 часов после инокуляции путем измерения активности FLuc в лизированных клетках (Cell Culture Lysis Reagent, Promega) с использованием коммерческого субстрата (Beetle-Juice, PJK) и планшетного люминометра (Hidex Sense Microplate Reader, Hidex). с программным обеспечением для считывания микропланшетов Hidex Sense (v0.5.41.0).
Серология
Мы определили серологию IgG SARS-CoV-2 с помощью количественного ИФА (домен шиповидного белка против SARS-CoV-2 S1/домен связывания рецептора (RBD) IgG SARS-CoV-2 QuantiVac, Euroimmun) в соответствии с инструкции производителя (разведение 1:400 или 1:600).Мы приводим уровни антител, выраженные в виде RU мл -1 , оцененные по калибровочной кривой, при этом значения выше 11 RU мл -1 определяются как положительные. Мы провели измерения домена шиповидного белка IgA против SARS-CoV-2 S1 или IgG против SARS-CoV-2 NCP в соответствии с инструкциями производителя (Euroimmun) и количества выраженных антител в виде соотношения IgA (оптическая плотность (OD), деленная на калибратор). Для анализа мы использовали программу AESKU.READER (AESKU.GROUP) и программное обеспечение Gen5 версии 2.01.
Для анализа мы использовали программу AESKU.READER (AESKU.GROUP) и программное обеспечение Gen5 версии 2.01.
sVNT для вариантов SARS-CoV-2
Для определения нейтрализующих антител против уханьского шипа вирус B.1.1.7-шип (Альфа), P.1-шип (B.1.1.28.1; Гамма) и B.1.351-шип (Бета) варианты SARS-CoV-2-S в плазме, мы недавно модифицировали нашу установлен сВНТ 19 . В этом анализе растворимый рецептор SARS-CoV-2 — ангиотензинпревращающий фермент 2 (ACE2) — связывается с 96-луночными планшетами, с которыми могут связываться различные очищенные меченые RBD шиповидного белка SARS-CoV-2. добавлено в анализ. Связывание дополнительно выявляют с помощью антител, меченных пероксидазой, и колориметрического количественного определения.Предварительная инкубация шиповидного белка с сывороткой или плазмой выздоравливающих пациентов или вакцинированных предотвращает последующее связывание с ACE2 в различной степени, в зависимости от количества присутствующих нейтрализующих антител. В частности, планшеты Maxisorp 96F (Nunc) покрывали рекомбинантным растворимым белком hACE2-Fc(IgG1) в количестве 300 нг на лунку в 50 мкл буфера для покрытия (30 мМ Na 2 CO 3 , 70 мМ NaHCO 3 , pH 9,6) при 4 °C в течение ночи. После блокировки hACE2-Fc(IgG1) планшеты промывали PBS, 0.05% Tween-20 (PBST) и блокировали разбавителем для анализа BD OptEIA на 1,5 часа при 37 °C. Тем временем образцы плазмы серийно разбавляли втрое, начиная с 1:20, а затем предварительно инкубировали в течение 1 часа при 37°C с 1,5 нг рекомбинантного шипа SARS-CoV-2 RBD штамма Wuhan (Trenzyme), Вариант B.1.1.7 (N501Y; Alpha), вариант B.1.351 (K417N, E484K, N501Y; Beta) или вариант P.1 (K417T, E484K, N501Y; Gamma) (последние три от SinoBiological), все с С-концевой His-меткой. BD OptEIA Assay Diluent использовался для приготовления образцов плазмы, а также разведений RBD.После предварительной инкубации с RBD с шипами SARS-CoV-2 образцы плазмы помещали на планшеты Maxisorp ELISA, покрытые hACE2, на 1 час при 37 °C.
В частности, планшеты Maxisorp 96F (Nunc) покрывали рекомбинантным растворимым белком hACE2-Fc(IgG1) в количестве 300 нг на лунку в 50 мкл буфера для покрытия (30 мМ Na 2 CO 3 , 70 мМ NaHCO 3 , pH 9,6) при 4 °C в течение ночи. После блокировки hACE2-Fc(IgG1) планшеты промывали PBS, 0.05% Tween-20 (PBST) и блокировали разбавителем для анализа BD OptEIA на 1,5 часа при 37 °C. Тем временем образцы плазмы серийно разбавляли втрое, начиная с 1:20, а затем предварительно инкубировали в течение 1 часа при 37°C с 1,5 нг рекомбинантного шипа SARS-CoV-2 RBD штамма Wuhan (Trenzyme), Вариант B.1.1.7 (N501Y; Alpha), вариант B.1.351 (K417N, E484K, N501Y; Beta) или вариант P.1 (K417T, E484K, N501Y; Gamma) (последние три от SinoBiological), все с С-концевой His-меткой. BD OptEIA Assay Diluent использовался для приготовления образцов плазмы, а также разведений RBD.После предварительной инкубации с RBD с шипами SARS-CoV-2 образцы плазмы помещали на планшеты Maxisorp ELISA, покрытые hACE2, на 1 час при 37 °C. RBD с шипами SARS-CoV-2, предварительно инкубированные с буфером, служили только в качестве отрицательного контроля для ингибирования. Планшеты трижды промывали PBST и инкубировали с конъюгированным с пероксидазой хрена (HRP) антителом против His-метки (клон HIS 3D5, предоставленный Helmholtz Zentrum München) в течение 1 ч при 37°C. Несвязанное антитело удаляли шестью промывками PBST. Колориметрический сигнал был получен при ферментативной реакции HRP с хромогенным субстратом 3,3′,5,5′-тетраметилбензидином (набор реагентов субстрата BD OptEIA TMB).Для остановки реакции добавляли равный объем 0,2 М H 2 SO 4 и измеряли оптическую плотность при 450 нм и 570 нм с помощью устройства для считывания микропланшетов SpectraMax iD3 (Molecular Devices) с использованием программного обеспечения SoftMax Pro версии 7.03. Для каждой лунки процент ингибирования рассчитывали из значений ОП после вычитания фоновых значений следующим образом: Ингибирование (%) = (1 - Значение ОП образца/Среднее значение ОП SARS-CoV-2 S RBD) × 100.
RBD с шипами SARS-CoV-2, предварительно инкубированные с буфером, служили только в качестве отрицательного контроля для ингибирования. Планшеты трижды промывали PBST и инкубировали с конъюгированным с пероксидазой хрена (HRP) антителом против His-метки (клон HIS 3D5, предоставленный Helmholtz Zentrum München) в течение 1 ч при 37°C. Несвязанное антитело удаляли шестью промывками PBST. Колориметрический сигнал был получен при ферментативной реакции HRP с хромогенным субстратом 3,3′,5,5′-тетраметилбензидином (набор реагентов субстрата BD OptEIA TMB).Для остановки реакции добавляли равный объем 0,2 М H 2 SO 4 и измеряли оптическую плотность при 450 нм и 570 нм с помощью устройства для считывания микропланшетов SpectraMax iD3 (Molecular Devices) с использованием программного обеспечения SoftMax Pro версии 7.03. Для каждой лунки процент ингибирования рассчитывали из значений ОП после вычитания фоновых значений следующим образом: Ингибирование (%) = (1 - Значение ОП образца/Среднее значение ОП SARS-CoV-2 S RBD) × 100.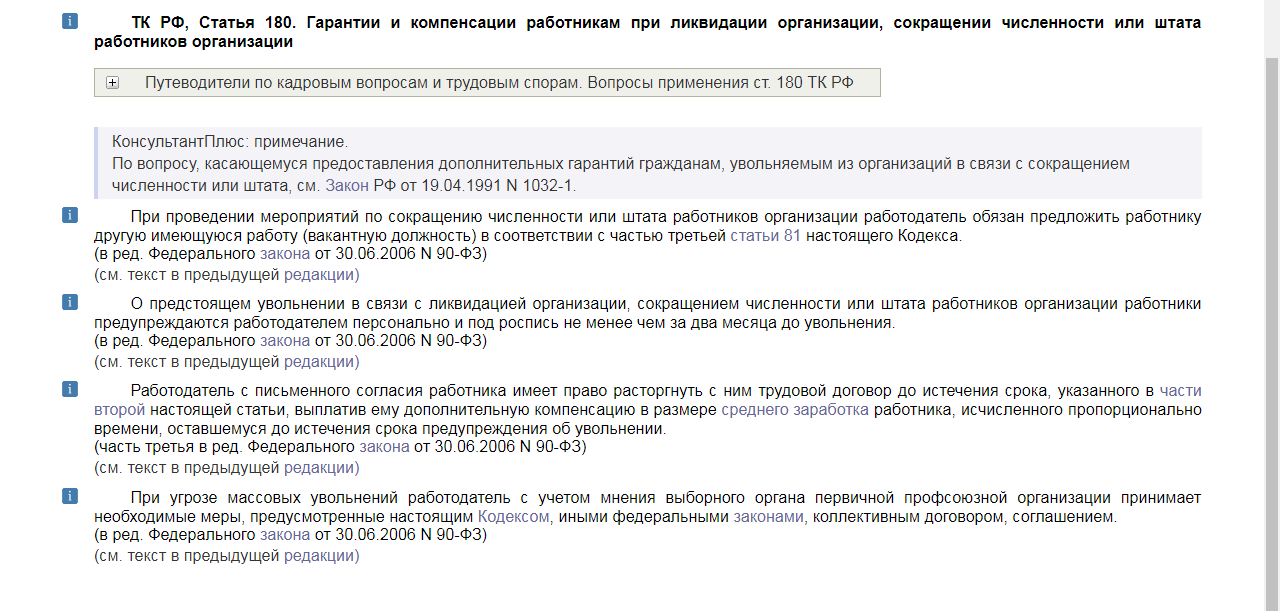 Определяли титры нейтрализующих sVNT. как разведение с уменьшением связывания > среднее + 2 с.д. значений из пула плазмы, состоящего из трех образцов плазмы до пандемии.
Определяли титры нейтрализующих sVNT. как разведение с уменьшением связывания > среднее + 2 с.д. значений из пула плазмы, состоящего из трех образцов плазмы до пандемии.
Белковые пептидные пулы SARS-CoV-2
Мы заказали 15 аминокислотных (аа) длинных и 10 аа-перекрывающихся пептидных пулов, охватывающих всю длину SARS-CoV2-Spike (-S) (всего 253 пептида), — Мембрана (-M) (всего 43 пептида), -нуклеокапсид (-N) (всего 82 пептида) или -оболочка (-E) (всего 12 пептидов; пептид № 4 не удалось синтезировать) из GenScript. Все лиофилизированные пептиды были синтезированы с чистотой более 95% и восстановлены в исходной концентрации 50 мг мл -1 в ДМСО (Sigma-Aldrich), за исключением девяти перекрывающихся пептидов SARS-CoV2-S (ном.24, 190, 191, 225, 226, 234, 244, 245 и 246), два для SARS-CoV2-M (№ 15 и 16), один для SARS-CoV2-N (№ 61) и все 12 SARS Пептиды -CoV2-E, растворенные в концентрации 25 мг/мл -1 из-за проблем с растворимостью. Все пептиды в ДМСО хранили до использования при температуре -80 °C.
Анализ повторной стимуляции Т-клеток
РВМС, выделенные с использованием градиента фиколла, ресуспендировали в концентрации 20 × 10 6 клеток на мл в полной среде RPMI (RPMI 1640 (Gibco)) с добавлением 10% FBS ( GE Healthcare Life Sciences), 1 мМ пирувата натрия, 50 мкМ β-меркаптоэтанола и 1% стрептомицина-пенициллина (все Gibco).Для стимуляции клетки разводили равными объемами пулов пептидов, содержащих S-белок или смесь M-, N- и E-белков. Пулы пептидов готовили в полной RPMI, содержащей брефельдин А (Sigma-Aldrich) в конечной концентрации 10 мкг/мл -1 . В конечной смеси каждый пептид имел концентрацию 2 мкг (~1,2 нмоль) мл -1 , за исключением пептидов SARS-CoV2-S 24, 190, 191, 225, 226, 234, 244, 245 и 246, Пептиды SARS-CoV2-M 15 и 16 и пептид SARS-CoV2-N 61, которые использовали в конечной концентрации 1 мкг мл -1 из-за проблем с растворимостью.В качестве отрицательного контроля мы стимулировали клетки ДМСО, используемым в максимальном объеме, соответствующем количеству ДМСО в пулах пептидов (равному 5% ДМСО в конечном объеме среды). В каждом эксперименте использовали клетки, стимулированные форбол-12-миристат-13-ацетатом (Calbiochem) и иономицином (Invitrogen) в конечной концентрации 50 нг мл -1 и 1500 нг мл -1 соответственно. внутренний положительный контроль. Затем клетки инкубировали в течение 12–16 часов при 37 °C, 5% CO 2 . После промывки клетки ресуспендировали в буфере MACS (PBS с добавлением 3% FBS и 2 мМ ЭДТА).Неспецифическое связывание антител блокировали путем инкубации образцов с 10% сывороткой мыши при 4°С в течение 15 мин. Далее, без отмывки, смесь антител анти-CD3-AF532 (UCHT1; № 58-0038-42, лот № 2288218; Invitrogen; 1:50), анти-CD4-BUV563 (RPA-T4; № 741353). , лот № 9333607; BD Biosciences; 1:200), анти-CD8-SparkBlue 550 (SK1; № 344760, лот № B326454; BioLegend; 1:200), анти-CD45RA (HI100; № 740298, лот № 0295003; BD Biosciences; 1:200), анти-CCR7 (G043H7; № 353230, № лота B335328; BioLegend; 1:50), анти-CD38 PerCP-eF710 (HB7; №46-0388-42, № лота 2044748; Инвитроген; 1:100) и Zombie NIR Fixable Viability Kit (№ 423106, № лота B323372; BioLegend).
В каждом эксперименте использовали клетки, стимулированные форбол-12-миристат-13-ацетатом (Calbiochem) и иономицином (Invitrogen) в конечной концентрации 50 нг мл -1 и 1500 нг мл -1 соответственно. внутренний положительный контроль. Затем клетки инкубировали в течение 12–16 часов при 37 °C, 5% CO 2 . После промывки клетки ресуспендировали в буфере MACS (PBS с добавлением 3% FBS и 2 мМ ЭДТА).Неспецифическое связывание антител блокировали путем инкубации образцов с 10% сывороткой мыши при 4°С в течение 15 мин. Далее, без отмывки, смесь антител анти-CD3-AF532 (UCHT1; № 58-0038-42, лот № 2288218; Invitrogen; 1:50), анти-CD4-BUV563 (RPA-T4; № 741353). , лот № 9333607; BD Biosciences; 1:200), анти-CD8-SparkBlue 550 (SK1; № 344760, лот № B326454; BioLegend; 1:200), анти-CD45RA (HI100; № 740298, лот № 0295003; BD Biosciences; 1:200), анти-CCR7 (G043H7; № 353230, № лота B335328; BioLegend; 1:50), анти-CD38 PerCP-eF710 (HB7; №46-0388-42, № лота 2044748; Инвитроген; 1:100) и Zombie NIR Fixable Viability Kit (№ 423106, № лота B323372; BioLegend). После окрашивания в течение 20 мин при комнатной температуре клетки промывали перед их фиксацией и пермеабилизацией (№ 554714, BD Biosciences) в соответствии с протоколом производителя. Далее внутриклеточные цитокины окрашивали анти-IFN-PE-Cy7 (B27; № 506518, лот № B326674; BioLegend; 1:100), анти-TNF-AF700 (Mab11; № 502928, лот № B326186; BioLegend; 1:50) и анти-IL-17A-BV421 (BL168; №.512322, № лота. Б317903; БиоЛегенда; 1:50) в течение 45 мин при комнатной температуре. Избыток антител смывали, а затем клетки собирали на спектральном проточном цитометре Cytek Aurora (Cytek), оснащенном пятью лазерами, работающими на длинах волн 355 нм, 405 нм, 488 нм, 561 нм и 640 нм. Все данные проточной цитометрии были получены с использованием SpectroFlo версии 2.2.0 (Cytek) и проанализированы с помощью FCS Express 7 (Denovo).
После окрашивания в течение 20 мин при комнатной температуре клетки промывали перед их фиксацией и пермеабилизацией (№ 554714, BD Biosciences) в соответствии с протоколом производителя. Далее внутриклеточные цитокины окрашивали анти-IFN-PE-Cy7 (B27; № 506518, лот № B326674; BioLegend; 1:100), анти-TNF-AF700 (Mab11; № 502928, лот № B326186; BioLegend; 1:50) и анти-IL-17A-BV421 (BL168; №.512322, № лота. Б317903; БиоЛегенда; 1:50) в течение 45 мин при комнатной температуре. Избыток антител смывали, а затем клетки собирали на спектральном проточном цитометре Cytek Aurora (Cytek), оснащенном пятью лазерами, работающими на длинах волн 355 нм, 405 нм, 488 нм, 561 нм и 640 нм. Все данные проточной цитометрии были получены с использованием SpectroFlo версии 2.2.0 (Cytek) и проанализированы с помощью FCS Express 7 (Denovo).
Проточный цитометрический анализ В-клеток, специфичных для шипа
Суммарные лейкоциты выделяли из цельной крови методом эритролиза в 0.83% раствор хлорида аммония. Затем изолированные клетки промывали, подсчитывали и ресуспендировали в PBS и окрашивали в течение 20 минут при комнатной температуре смесью антител, содержащей антитела, перечисленные на рис. 4 с расширенными данными, вместе с белком шип-mNEONGreen (5 мкг на реакцию; получение будет описано в другом месте). . После одной промывки образцы собирали на спектральном проточном цитометре и данные анализировали, как описано выше.
Затем изолированные клетки промывали, подсчитывали и ресуспендировали в PBS и окрашивали в течение 20 минут при комнатной температуре смесью антител, содержащей антитела, перечисленные на рис. 4 с расширенными данными, вместе с белком шип-mNEONGreen (5 мкг на реакцию; получение будет описано в другом месте). . После одной промывки образцы собирали на спектральном проточном цитометре и данные анализировали, как описано выше.
Количественная оценка высвобождения IFN-γ и TNF-α
Далее, 0.5 мл цельной крови стимулировали выбранными производителем частями домена S1 шиповидного белка SARS-CoV-2 в течение 20–24 часов. Мы провели отрицательный и положительный контроль в соответствии с инструкциями производителя (анализ высвобождения гамма-интерферона SARS-CoV-2 (Euroimmun)). После стимуляции супернатанты собирали после центрифугирования и исследовали с помощью набора LEGENDplex (BioLegend) в соответствии с инструкциями производителя. Данные повторных измерений были получены с помощью проточного цитометра LSR II (BD Biosciences) с использованием BD FACSDiva версии 8. 0.1 и проанализированы с помощью LEGENDplex Data Analysis Software Suite, Gen5, версия 2.01.
0.1 и проанализированы с помощью LEGENDplex Data Analysis Software Suite, Gen5, версия 2.01.
Статистика
Статистический анализ был выполнен с использованием GraphPad Prism версии 8.4 (программное обеспечение GraphPad) и SPSS версии 20.0.0 (IBM, SPSS Statistics). Для сравнения уровней спайк-специфических IgG и IgA, а также для сравнения процентного содержания цитокин-секретирующих Т-клеток или концентрации цитокинов в сыворотке мы использовали парный тест t (внутри групп) или двусторонний дисперсионный анализ. (ANOVA) с последующим тестом множественных сравнений Сидака (между группами).Для сравнения титров sVNT мы использовали критерий хи-квадрат для тренда. Различия считались значимыми, если P < 0,05. Корреляцию между значениями sVNT и pVNT рассчитывали с использованием однократного линейного регрессионного анализа.
Утверждение комитета по этике
Исследование CoCo и анализ, проведенный для этой статьи, были одобрены внутренним наблюдательным советом Ганноверской медицинской школы (институциональный наблюдательный совет № 8973_BO-K_2020, поправка от декабря 2020 г. ).
).
Сводка отчетов
Дополнительная информация о дизайне исследования доступна в Сводке отчетов по исследованиям природы, связанной с этой статьей.
Текущие применения радиочастотной абляции в онкологии
Abstract
Радиочастотная абляция (РЧА) использует высокочастотный переменный ток для нагревания объема ткани вокруг игольчатого электрода, чтобы вызвать очаговый коагуляционный некроз с минимальным повреждением окружающих тканей. РЧА может выполняться через открытым, лапароскопическим или чрескожным доступом под визуальным контролем и под общей или местной анестезией. Достижения в области механизмов доставки, конструкций электродов и генераторов более высокой мощности увеличили максимальный объем, который можно аблировать, при этом максимизировав онкологические результаты.Как правило, РЧА используется для контроля локального роста опухоли, предотвращения рецидивов, облегчения симптомов и улучшения выживаемости у подгруппы пациентов, которые не являются кандидатами на хирургическую резекцию. Его эквивалентность хирургической резекции еще предстоит доказать в крупных рандомизированных контролируемых исследованиях. В настоящее время использование РЧА хорошо описано в качестве основного или адъювантного метода лечения ограниченной, но нерезектабельной гепатоцеллюлярной карциномы, метастазов в печени, особенно метастазов колоректального рака, первичных опухолей легких, почечно-клеточного рака, костных метастазов и остеоид-остеом.Роль РЧА в первичной терапии рака молочной железы на ранних стадиях все еще развивается. В этом обзоре обсуждаются общие особенности РЧА и описывается ее роль при часто встречающихся солидных опухолях.
Его эквивалентность хирургической резекции еще предстоит доказать в крупных рандомизированных контролируемых исследованиях. В настоящее время использование РЧА хорошо описано в качестве основного или адъювантного метода лечения ограниченной, но нерезектабельной гепатоцеллюлярной карциномы, метастазов в печени, особенно метастазов колоректального рака, первичных опухолей легких, почечно-клеточного рака, костных метастазов и остеоид-остеом.Роль РЧА в первичной терапии рака молочной железы на ранних стадиях все еще развивается. В этом обзоре обсуждаются общие особенности РЧА и описывается ее роль при часто встречающихся солидных опухолях.
Ключевые слова: Радиочастотная абляция, Гепатоцеллюлярная карцинома, Колоректальный рак с метастазами в печень, Рак легких, Почечно-клеточная карцинома
Основной наконечник: использование и его эквивалентность или отсутствие эквивалентности хирургической резекции.Мы выделили исследования, в которых сообщалось об отдаленных онкологических исходах, связанных с применением РЧА при первичных и метастатических опухолях печени и легких, и описывалась развивающаяся роль РЧА при опухолях молочной железы и солидных почечных опухолях.
ВВЕДЕНИЕ
Хирургическая резекция всех злокачественных клеток остается золотым стандартом лечения большинства солидных опухолей[1]. Однако хирургическая резекция не всегда возможна у пациентов с сопутствующими заболеваниями или плохим функциональным статусом, когда резекция может быть связана с высокой заболеваемостью и смертностью.В результате были разработаны различные методы местной абляции, в том числе химические (этанол, уксусная кислота, горячий физиологический раствор) и термические (радиочастотная абляция, микроволновая абляция, лазерная абляция, криоабляция) для уничтожения раковых клеток in situ . Радиочастотная абляция (РЧА) вышла на передний план среди этих местных абляционных методов благодаря совершенствованию технологии, которая максимизирует эффективность и простоту использования при минимизации сопутствующих осложнений. В настоящее время РЧА используется как в лечебном, так и в паллиативном лечении солидных опухолей по всему телу.Эта минимально инвазивная методика может служить как в качестве лечения пациентов, не являющихся кандидатами на хирургическое вмешательство, так и в качестве дополнения к хирургическому вмешательству, облегчающему резекцию или в сочетании с хирургическим вмешательством для достижения полного контроля над опухолевой массой.
ТЕХНИЧЕСКИЕ ХАРАКТЕРИСТИКИ РЧА
РЧА использует радиоволны низкой частоты (460–480 кГц) и большой длины волны для выработки тепла внутри опухолевой массы, вызывающего термический коагуляционный некроз. РЧА отличается от других местных методов тем, что сам электрод не подает тепло.Игольчатые электроды подают переменный электрический ток, который проходит от электрода к заземляющей площадке (монополярный) или между двумя электродами (биполярный). Поскольку ионы в ткани пытаются следовать переменному пути тока, ионное возбуждение создает тепло от трения. Это трение нагревает окружающие ткани до 50-100 °C, вызывая мгновенный коагуляционный некроз. Температуры выше 100 °C приводят к высыханию тканей и зарядке с потерей ионов, что приводит к прекращению тока.Это приводит к резкому повышению импеданса [2], что ограничивает объем ткани, которая может быть успешно удалена.
Энергия кончика электрода создает температуру, пропорциональную квадрату радиочастотного тока, которая, в свою очередь, уменьшается пропорционально квадрату диаметра электрода[2]. Опухоли большего размера требуют перекрывающихся сфер, что увеличивает риск неполного некроза и, следовательно, местного рецидива. За последние несколько лет достижения в области механизмов доставки, которые могут либо увеличить количество депонируемой энергии, либо проводимость тепла через ткань, увеличили область ткани, которая может быть подвергнута абляции [3].В настоящее время пять компаний производят коммерчески доступные системы РЧА, четыре из которых одобрены Управлением по санитарному надзору за качеством пищевых продуктов и медикаментов и доступны в США[4]. Технические характеристики каждой системы представлены в таблице.
Опухоли большего размера требуют перекрывающихся сфер, что увеличивает риск неполного некроза и, следовательно, местного рецидива. За последние несколько лет достижения в области механизмов доставки, которые могут либо увеличить количество депонируемой энергии, либо проводимость тепла через ткань, увеличили область ткани, которая может быть подвергнута абляции [3].В настоящее время пять компаний производят коммерчески доступные системы РЧА, четыре из которых одобрены Управлением по санитарному надзору за качеством пищевых продуктов и медикаментов и доступны в США[4]. Технические характеристики каждой системы представлены в таблице.
Таблица 1
Радиочастотные системы абляции коммерчески доступны в Соединенных Штатах [4,8] Многозондовые матричные электроды, в которых несколько зубцов одновременно подают ток, обеспечивают зоны коагуляции 3-5 см. Было разработано несколько стратегий для снижения толерантности опухоли к теплу и повышения эффективности методов термоабляции. Эффект «теплоотвода», создаваемый близостью опухолей к крупным сосудам, которые могут рассеивать тепло, является основным механизмом, с помощью которого можно ограничить степень термического повреждения [5, 6]. РЧА может выполняться чрескожно, во время лапароскопической или открытой операции. У каждого есть свои преимущества и недостатки, и подход будет зависеть от состояния пациента, характеристик опухоли, таких как расположение, размер, количество и характер роста, а также опыта и предпочтений поставщика [8].На сегодняшний день недостаточно данных, указывающих, какой метод родоразрешения является предпочтительным, из-за отсутствия рандомизированных контрольных исследований и различий в характеристиках пациентов и опухолей между исследованиями с использованием одной методики. Преимущество чрескожного доступа заключается в том, что он выполняется в сознании или под глубокой седацией, что дает возможность пациентам с более высоким хирургическим риском. Обычно это можно сделать амбулаторно или с очень коротким пребыванием в больнице, и при необходимости можно проводить несколько раз. Чрескожный доступ также может выполняться под анестезией. Другими преимуществами этого метода являются использование сонографии, компьютерной томографии (КТ) или магнитно-резонансной томографии (МРТ) для точного размещения электродов.В тех же условиях во время процедуры можно провести УЗИ с контрастным усилением или КТ с контрастным усилением, чтобы проверить адекватность абляции. Открытая РЧА обеспечивает лучшую визуализацию и возможность манипулировать соседними структурами. Его преимущество заключается в возможности обнаружения скрытых метастазов с использованием интраоперационного ультразвука (УЗИ) и позволяет проводить лечение в более широком анатомическом диапазоне. Другим преимуществом РЧА печени является возможность окклюзии портального притока (маневр Прингла), что, как описано выше, снижает теплоотдачу и, следовательно, увеличивает объем аблируемой ткани. Этот метод особенно ценен, когда опухоли расположены в непосредственной близости от сосудистых структур. Лапароскопическая РЧА сочетает в себе многие преимущества как чрескожного, так и открытого доступов. Он минимально инвазивен с меньшей болезненностью большого разреза, но при этом позволяет лучше визуализировать опухоль и соседние структуры для оптимизации стадирования. Пневмоперитонеум также может работать аналогично маневру Прингла и уменьшать эффект теплоотвода в опухолях вблизи крупных сосудов за счет уменьшения портального кровотока [11]. Он также позволяет выполнять резекцию или смещение структур, прилегающих к опухолям, что невозможно выполнить с помощью чрескожной техники. Визуализация играет важную роль в диагностике и локализации опухоли, в мониторинге зоны абляции в режиме реального времени, в оценке реакции ткани на радиочастотную терапию и, наконец, в последующем наблюдении за пациентом. РЧ-датчик обычно находится под контролем КТ или УЗИ, а процедура РЧА контролируется с помощью УЗИ в реальном времени. Зоны абляции видны на УЗИ как гиперэхогенные области, которые представляют собой микропузырьки, образующиеся в результате испарения интерстициальной жидкости из аблированной ткани. Целью РЧА обычно является удаление 1 см границы нормальной ткани, окружающей опухоль со всех сторон [8,14,15]. Этот хирургический край необходим из-за трудности точного определения протяженности зоны коагуляции и из-за возможности микроскопического злокачественного новообразования, окружающего макроскопическую опухоль [8]. РЧА является относительно безопасной процедурой со смертностью от 0,3% до 0,8% и заболеваемостью от 2% до 10%[16,17]. Осложнения включают постпроцедурную боль, синдром после РЧА с лихорадкой и гриппоподобными симптомами, которые обычно проходят в течение первых 24 часов, ожоги кожи из-за неправильно размещенных заземляющих пластин, термическое повреждение соседних структур, кровотечение, вторичную инфекцию и образование опухоли. этого можно избежать, прижигая ход иглы при извлечении зонда [8]. Наиболее обширная литература по РЧА для лечения солидных опухолей посвящена ее использованию при злокачественных новообразованиях печени, как первичных, так и метастатических. Большая часть литературы, посвященной метастазам в печень, получена в результате одногрупповых, ретроспективных или проспективных исследований, оценивающих РЧА для лечения нерезектабельных колоректальных метастазов. В таких исследованиях резекция печени превосходит как одну РЧА, так и комбинацию РЧА с резекцией печени в отношении местного рецидива и общей выживаемости [26]. Однако при открытой резекции могут быть обнаружены дополнительные опухоли на поверхности печени или глубокие метастазы при интраоперационном УЗИ.Эти дополнительные поражения можно резецировать или лечить с помощью интраоперационной РЧА. Существует значительная перекрывающаяся вариабельность 5-летней выживаемости и частоты местных рецидивов из-за различий в определении местного рецидива, критериях включения в отношении нерезектабельности, степени внепеченочного поражения и характеристиках пациентов и опухолей между исследованиями. Частота местных рецидивов варьировала от 9% до 40%, а общая 5-летняя выживаемость варьировала от 18% до 30% (таблица).Наилучшие результаты были у пациентов с солитарными опухолями менее 3 см и несколько хуже у пациентов с 3 и менее опухолями менее 3 см [27]. Местный рецидив был значительно выше у пациентов с опухолями размером от 3 до 5 см [20]. Ретроспективные исследования, сравнивающие резекцию печени с РЧА у пациентов, потенциально операбельных, но не являющихся кандидатами на операцию из-за сопутствующих заболеваний или отказа, продемонстрировали снижение местного рецидива и улучшение общей выживаемости при резекции печени [28]. Исследования, сообщающие о выживаемости после использования радиочастотной абляции метастазов колоректального рака в печень РЧА все чаще применяется к злокачественным узлам в легких для локального контроля, а также для паллиативного лечения, так как его осуществимость и эффективность все больше упоминаются в литературе. В настоящее время недостаточно доказательств того, что РЧА сравнима с хирургической резекцией.В настоящее время нет проспективных рандомизированных контролируемых исследований, сравнивающих РЧА со стандартными вариантами хирургического лечения пациентов со злокачественными узлами в легких. Данные ограничены сериями случаев с различиями в количестве первичных и вторичных поражений легких, критериях нерезектабельности, количестве предыдущих резекций, истории предшествующей лучевой терапии, различиях в протоколах наблюдения и критериях определения степени ответа на лечение РЧА. Однако небольшая серия сопоставимых случаев из 22 пациентов, сравнивающих РЧА с резекцией у пациентов с немелкоклеточным раком легкого (НМРЛ) стадии I , продемонстрировала сопоставимую выживаемость у пациентов с РЧА через 1, 2 и 5 лет [30].Исследование RAPTURE, крупное проспективное многоцентровое исследование с одной группой, с использованием РЧА у пациентов с ранней стадией НМРЛ или метастазами в легкие, продемонстрировало 1-летнюю и 2-летнюю общую выживаемость 70% и 48% соответственно у пациентов с первичными опухолями легких и 89% и 66. % 1-летняя и 2-летняя общая выживаемость у больных с колоректальными метастазами. Онкоспецифическая выживаемость была выше в обеих группах; 92% и 73% через 1 и 2 года в когорте НМРЛ и 91% и 68% в когорте с колоректальными метастазами. 1-, 2-, 3-летняя общая выживаемость пациентов с ранними стадиями первичного рака легкого, получавших РЧА, варьирует от 70 до 90%, от 48 до 84%, от 25 до 74% соответственно (таблица).Это сравнимо с 1, 3, 5-летней общей выживаемостью пациентов, перенесших лобэктомию или сегментарную резекцию по поводу рака легкого на ранней стадии [31-34]. Исследования выживаемости с использованием радиочастотной абляции первичных опухолей легких и метастазов Медиана заболеваемости и смертности, связанных с процедурой, составляет 37. Роль РЧА при раке молочной железы все еще выясняется. Существует растущая тенденция к методам сохранения груди, которые минимизируют рубцы, деформацию груди и улучшают общий косметический эффект после процедуры.Несколько небольших исследований в одном учреждении установили осуществимость РЧА и выявили возможные осложнения (таблица). В большинстве этих исследований за РЧА следовала лампэктомия или мастэктомия либо немедленно, либо отсроченно. Процедура проводилась под местной или общей анестезией в зависимости от того, была ли резекция отсроченной или сразу после РЧА соответственно. Исследования выживаемости с использованием радиочастотной абляции первичных опухолей легких и метастазов Критерии отбора пациентов были строгими, включая в основном пациентов с инвазивными опухолями размером менее 2 см; в нескольких исследованиях была небольшая часть пациентов с неинвазивными опухолями.Наличие обширного внутрипротокового компонента также было относительным противопоказанием к РЧА. В настоящее время нет исследований, сравнивающих РЧА с хирургической резекцией, а также долгосрочных исследований, отражающих частоту местных рецидивов или выживаемость у пациентов, получающих РЧА вместо хирургической резекции. Реакция пациента на РЧА была благоприятной. Oura и коллеги [41] ретроспективно оценили косметические результаты, которые оказались отличными у 43 пациентов (83%), хорошими у 6 пациентов (12%) и удовлетворительными у 3 пациентов (6%). Авторы установили, что основным фактором, приводящим к ухудшению косметических свойств, является образование новообразований в месте РЧА, особенно у женщин с маленькой грудью.Это также может привести к повышенной тревожности пациента. Прогресс в применении РЧА для опухолей молочной железы в настоящее время сдерживается нашей способностью точно оценивать состояние края, которое является критическим фактором в частоте местных рецидивов. Как и при других солидных опухолях, РЧА все чаще применяется для лечения опухолей почек, поскольку менее инвазивные и нефронсберегающие методы, включая частичную нефрэктомию и лапароскопическую нефрэктомию, доказали свою сопоставимую 5-летнюю безрецидивность выживания[42]. В настоящее время РЧА в качестве основного лечения злокачественных новообразований почки ограничена в исследованиях избранной группой пациентов с ранним заболеванием T1a или у которых хирургическая резекция невозможна. К ним относятся пациенты только с одной почкой, многоочаговым заболеванием фон Хиппеля Линдау, ограниченной функцией почек, пожилые пациенты или пациенты с сопутствующими заболеваниями, которые являются плохими кандидатами на операцию [8, 43-45]. Исследования выживаемости после радиочастотной абляции солидных опухолей почки 94 P, 2 L 9 290 90 2994 Эффективность РЧА для лечения остеоид-остеом давно доказана. Совсем недавно РЧА стали применять в качестве паллиативного метода лечения болезненных метастатических поражений костей. Внешнее лучевое облучение остается золотым стандартом лечения локализованной боли в костях из метастатического очага. Однако 20-30% пациентов не реагируют на фармакотерапию и сопротивляются ей [51,52]. Кроме того, пациенты, ранее облученные в рецидивирующем очаге, могут не иметь права на повторную лучевую терапию. 1. 2. Ауригемма Г.П.,
Гааш У.Х.
Клиническая практика. Диастолическая сердечная недостаточность. N Английский J Med .
2004;351(11):1097–1105. 3. Янси CW,
Джессап М,
Бозкурт Б,
и другие.
Руководство ACCF/AHA 2013 года по лечению сердечной недостаточности: отчет Целевой группы Фонда Американского колледжа кардиологов/Американской кардиологической ассоциации по практическим рекомендациям. J Am Coll Cardiol .
2013;62(16):e147–e239. 4. Ли Д.С.,
Гона П,
Васан Р.С.,
и другие.
Связь патогенеза заболевания и факторов риска с сердечной недостаточностью с сохраненной или сниженной фракцией выброса: результаты исследования сердца Framingham, проведенного Национальным институтом сердца, легких и крови. 5. Пониковоски П,
Воорс А.А.,
Анкер С.Д.,
и другие.;
Целевая группа по диагностике и лечению острой и хронической сердечной недостаточности Европейского общества кардиологов (ESC).
Руководство ESC 2016 г. по диагностике и лечению острой и хронической сердечной недостаточности. Сердечная недостаточность Eur J .
2016;18(8):891–975. 6. Мадхок В.,
Фальк Г,
Роджерс А,
Стразерс А.Д.,
Салливан FM,
Фэйи Т.
Точность симптомов, признаков и диагностических тестов при диагностике дисфункции левого желудочка в первичной медико-санитарной помощи: систематический обзор диагностической точности. BMC Fam Pract .
2008; 9:56. 7. Мант Дж.,
Дуст Дж,
Роальф А,
и другие.
Систематический обзор и метаанализ данных отдельных пациентов о диагностике сердечной недостаточности с моделированием последствий различных диагностических стратегий в первичной помощи. 8. Мартиндейл Дж.Л.,
Вакаи А,
Коллинз СП,
и другие.
Диагностика острой сердечной недостаточности в отделении неотложной помощи: систематический обзор и метаанализ. Академия скорой медицинской помощи .
2016;23(3):223–242. 9. Келдер Дж.С.,
Коуи М.Р.,
Макдонах Т.А.,
и другие.
Количественная оценка добавленной стоимости BNP при подозрении на сердечную недостаточность в общей практике: метаанализ данных отдельных пациентов. Сердце .
2011;97(12):959–963. 10. Роальф А.К.,
Мант Дж,
Дуст Дж.А.,
и другие.
Разработка и первоначальная валидация простого инструмента принятия клинических решений для прогнозирования наличия сердечной недостаточности в условиях первичной медико-санитарной помощи: правило MICE (мужчина, инфаркт, крепитация, отек). Сердечная недостаточность Eur J .
2012;14(9):1000–1008. 11. Фонсека С,
Оливейра АГ,
Мота Т,
и другие.;
Исследователи ЭПИКА.
Оценка эффективности и согласованности клинических опросников для диагностики сердечной недостаточности в условиях первичной медико-санитарной помощи. Сердечная недостаточность Eur J .
2004;6(6):813–820, 821–822. 12. Макки Пенсильвания,
Кастелли ВП,
Макнамара премьер-министр,
Каннель ВБ.
Естественная история застойной сердечной недостаточности: исследование Framingham. N Английский J Med .
1971; 285 (26): 1441–1446. 13. Келдер Дж.К.,
Крамер М.Дж.,
ван Вейнгаарден Дж.,
и другие.
Диагностическое значение физикального осмотра и дополнительных тестов у пациентов первичного звена с подозрением на сердечную недостаточность. Тираж .
2011;124(25):2865–2873. 14. Васан Р.С.,
Леви Д.
Определение диастолической сердечной недостаточности: призыв к стандартизированным диагностическим критериям. Тираж .2000;101(17):2118–2121. 15. Чиннаян К.М.,
Александр Д,
Мэдденс М,
Маккалоу П.А.
Учебная программа по кардиологии: комплексная диагностика и лечение диастолической сердечной недостаточности. Am Heart J .
2007;153(2):189–200. 16. Юсуф С.,
Пфеффер М.А.,
Сведберг К,
и другие.;
CHARM Следователи и комитеты.
Эффекты кандесартана у пациентов с хронической сердечной недостаточностью и сохраненной фракцией выброса левого желудочка: исследование CHARM-Preserved. Ланцет .
2003;362(9386):777–781. 17. Масси Б.М.,
Карсон ЧП,
МакМюррей Джей Джей,
и другие.;
I-PRESERVE Следователи.
Ирбесартан у больных с сердечной недостаточностью и сохранной фракцией выброса. N Английский J Med .
2008;359(23):2456–2467. 18. Херан Б.С.,
Мусини В.М.,
Бассет К,
Тейлор Р.С.,
Райт Дж.М.
Блокаторы рецепторов ангиотензина при сердечной недостаточности Cochrane Database Syst Rev .2012;(4):CD003040. 19. Клеланд Дж. Г.,
Тендера М,
Адамус Дж,
Фримантл Н,
Полонский Л,
Тейлор Дж.;
Исследователи PEP-CHF.
Исследование периндоприл у пожилых людей с хронической сердечной недостаточностью (PEP-CHF). Европейское сердце J .
2006;27(19):2338–2345. 20. Флатер, доктор медицинских наук,
Шибата МС,
Пальто ЭйДжей,
и другие.;
СТАРШИЕ Следователи.
Рандомизированное исследование влияния небиволола на смертность и частоту госпитализаций по сердечно-сосудистым заболеваниям у пожилых пациентов с сердечной недостаточностью (SENIORS). Европейское сердце J .
2005;26(3):215–225. 21. ван Вельдхейзен Д.Дж.,
Коэн-Соляль А,
Бём М,
и другие.;
СТАРШИЕ Следователи.
Бета-блокада небивололом у пожилых пациентов с сердечной недостаточностью с нарушенной и сохраненной фракцией выброса левого желудочка: данные SENIORS (Изучение влияния вмешательства с небивололом на исходы и повторную госпитализацию у пожилых с сердечной недостаточностью). J Am Coll Cardiol .
2009;53(23):2150–2158. 22. Ямамото К.,
Оригаса Х,
Хори М;
Исследователи J-DHF.
Влияние карведилола на сердечную недостаточность с сохраненной фракцией выброса: японское исследование диастолической сердечной недостаточности (J-DHF). Сердечная недостаточность Eur J .
2013;15(1):110–118. 23. Эдельманн Ф.,
Вахтер Р,
Шмидт АГ,
и другие.;
Исследователи Альдо-ДХФ.
Влияние спиронолактона на диастолическую функцию и переносимость физической нагрузки у пациентов с сердечной недостаточностью с сохраненной фракцией выброса: рандомизированное контролируемое исследование Aldo-DHF. ДЖАМА .
2013;309(8):781–791. 24. Питт Б.,
Пфеффер М.А.,
Ассманн С.Ф.,
и другие.;
Исследователи TOPCAT.
Спиронолактон при сердечной недостаточности с сохраненной фракцией выброса. N Английский J Med .
2014;370(15):1383–1392. 25. Редфилд М.М.,
Анстром К.Дж.,
Левин Дж.А.,
и другие.;
Сеть клинических исследований сердечной недостаточности NHLBI.
Изосорбида мононитрат при сердечной недостаточности с сохраненной фракцией выброса. N Английский J Med .
2015;373(24):2314–2324. 26. Ахмед А.,
Богатый МВт,
Флег Дж.Л.,
и другие.
Влияние дигоксина на заболеваемость и смертность при диастолической сердечной недостаточности: исследование исследовательской группы вспомогательных средств наперстянки. Тираж .
2006;114(5):397–403. 27. Хашим Т.,
Эльбаз С,
Патель К,
и другие.
Дигоксин и 30-дневная госпитализация по любой причине у пожилых пациентов с хронической диастолической сердечной недостаточностью. Am J Med .
2014;127(2):132–139. 28. Редфилд М.М.,
Чен Х.Х.,
Борлауг Б.А.,
и другие.;
РАССЛАБЬТЕСЬ Пробная версия.
Влияние ингибирования фосфодиэстеразы-5 на толерантность к физической нагрузке и клиническое состояние при сердечной недостаточности с сохраненной фракцией выброса: рандомизированное клиническое исследование. ДЖАМА .
2013;309(12):1268–1277. 29. Фукута Х,
Гото Т,
Ваками К,
Охте Н.
Влияние лекарств и физических упражнений на функциональную способность и качество жизни при сердечной недостаточности с сохраненной фракцией выброса: метаанализ рандомизированных контролируемых исследований. Eur J Prev Cardiol .
2016;23(1):78–85. 30. Хван С.Дж.,
Меленовский В,
Борлауг Б.А.
Последствия ишемической болезни сердца при сердечной недостаточности с сохраненной фракцией выброса. J Am Coll Cardiol .
2014; 63 (25 пт А): 2817–2827. 31. Фарис РФ,
Флатер М,
Перселл Х,
Пул-Уилсон, Пенсильвания,
Пальто Эй Джей.
Диуретики при сердечной недостаточности Cochrane Database Syst Rev .
2012;(2):CD003838. 32. Кейн Г.К.,
Карон БЛ,
Махони Д.В.,
и другие.
Прогрессирование диастолической дисфункции левого желудочка и риск сердечной недостаточности. ДЖАМА .
2011;306(8):856–863. 33. Перес де Исла Л.,
Каньядас В,
Контрерас Л,
и другие.
Диастолическая сердечная недостаточность у пожилых: госпитальные и отдаленные результаты после первого эпизода. Int J Cardiol .
2009;134(2):265–270. 34.Гербер Ю,
Уэстон С.А.,
Редфилд ММ,
и другие.
Современная оценка эпидемии сердечной недостаточности в округе Олмстед, штат Миннесота, с 2000 по 2010 год. JAMA Intern Med .
2015;175(6):996–1004. 35. Трибуйой С,
Русинару Д,
Махджуб Х,
и другие.
Прогноз сердечной недостаточности с сохраненной фракцией выброса: 5-летнее проспективное популяционное исследование. Европейское сердце J .
2008;29(3):339–347. 36. Комайда М,
Карсон ЧП,
Хетцель С,
и другие.
Факторы, связанные с исходом при сердечной недостаточности с сохраненной фракцией выброса: результаты исследования ирбесартана при сердечной недостаточности с сохраненной фракцией выброса (I-PRESERVE). Сердечная недостаточность .
2011;4(1):27–35. 37. Берк М.А.,
Кац ДХ,
Бессинк Л,
и другие.
Прогностическое значение патофизиологических маркеров у больных с сердечной недостаточностью и сохранной фракцией выброса. Сердечная недостаточность .
2014;7(2):288–299. 38. Клеланд Дж. Г.,
Тейлор Дж,
Фримантл Н,
Гуд КМ,
Ригби А.С.,
Тендера М.
Взаимосвязь между концентрациями N-концевого промозгового натрийуретического пептида в плазме и характеристиками и исходом у пациентов с клиническим диагнозом диастолической сердечной недостаточности: отчет исследования PEP-CHF. Сердечная недостаточность Eur J .
2012;14(5):487–494. 39. Кинг М,
Кингери Дж,
Кейси Б.Диагностика и оценка сердечной недостаточности. Семейный врач .
2012;85(12):1161–1168. 40. Сатпати С,
Мишра ТК,
Сатпати Р,
Сатпати Гонконг,
Бароне Э.
Диагностика и лечение диастолической дисфункции и сердечной недостаточности [опубликованное исправление появляется в Am Fam Physician. 2008;78(4):434]. Семейный врач .
2006;73(5):841–846. 41. Гутьеррес К.,
Бланшар ДГ.
Диастолическая сердечная недостаточность: проблемы диагностики и лечения. Семейный врач .
2004;69(11):2609–2616. RFA System Электроды Генератор Power / частота Система управления алгоритм, используемый для максимального максимального объема Boston Scientific 14 манометр, 10-12 клетки, зонтик в форме 200 Вт / 460 кГц Коаксиальная система Коаксиальная система Valleylab 17.  5 калибра, одна охлаждаемая иглы или три охлажденных иглы в треугольном кластере
5 калибра, одна охлаждаемая иглы или три охлажденных иглы в треугольном кластере 200 Вт / 480 кГц Контрема Охлаждение RITA Медицинские системы 250 Вт / 460 кГц STARBURST XL 14 Датчик, 9 клетки, Рождественская елка Форма Максимальный диаметр 5 см STARBURST XLI 14 CAMENT, 9 TINES, MAX диаметр 7 см Starburst Flex 13 Датчик 9 Berchtold 9-14 датчик 60 Вт / 375 кГц Импеданс или контролируемый температурой на мокрный электроду  Электроды с внутренним охлаждением (или с холодным наконечником) также позволяют увеличить объем абляции. Хотя охлаждение электрода путем непрерывного введения жидкости в просвет кажется парадоксальным, такое охлаждение приводит к отсутствию локального обугливания вокруг неизолированного кончика электрода, что обеспечивает более длительный ток. Более продолжительный ток обеспечивает больший объем локальной коагуляции тканей по сравнению с электродами без внутреннего охлаждения. Влажные электроды с использованием физиологического раствора (изотонического или гипертонического), вводимого через электрод в окружающие ткани, повышают проводимость за счет введения большего количества ионов в ткань, увеличивая ток и, таким образом, обеспечивая более длительную продолжительность тока и увеличивая объем коагуляции.
Электроды с внутренним охлаждением (или с холодным наконечником) также позволяют увеличить объем абляции. Хотя охлаждение электрода путем непрерывного введения жидкости в просвет кажется парадоксальным, такое охлаждение приводит к отсутствию локального обугливания вокруг неизолированного кончика электрода, что обеспечивает более длительный ток. Более продолжительный ток обеспечивает больший объем локальной коагуляции тканей по сравнению с электродами без внутреннего охлаждения. Влажные электроды с использованием физиологического раствора (изотонического или гипертонического), вводимого через электрод в окружающие ткани, повышают проводимость за счет введения большего количества ионов в ткань, увеличивая ток и, таким образом, обеспечивая более длительную продолжительность тока и увеличивая объем коагуляции. Маневр Прингла, который включает перекрытие портального притока во время открытой РЧА. Было показано, что это улучшает объем коагуляции ткани (опухоли) за счет увеличения локального отложения тепла, а не рассеивания тепла в воротной вене [6,7].Также было показано, что повреждение тканей в результате химиотерапии и гипоксическое повреждение опухолевых клеток в результате эмболизации повышают чувствительность опухоли к гипертермии. Также был продемонстрирован синергетический эффект между неоадъювантной трансартериальной химиоэмболизацией и РЧА при лечении гепатоцеллюлярной карциномы [7].
Маневр Прингла, который включает перекрытие портального притока во время открытой РЧА. Было показано, что это улучшает объем коагуляции ткани (опухоли) за счет увеличения локального отложения тепла, а не рассеивания тепла в воротной вене [6,7].Также было показано, что повреждение тканей в результате химиотерапии и гипоксическое повреждение опухолевых клеток в результате эмболизации повышают чувствительность опухоли к гипертермии. Также был продемонстрирован синергетический эффект между неоадъювантной трансартериальной химиоэмболизацией и РЧА при лечении гепатоцеллюлярной карциномы [7]. Техника РЧА
 В исследовании, сравнивающем открытый, лапароскопический и чрескожный доступы к опухолям печени, не было различий в смертности, серьезных осложнениях или общей выживаемости; но открытый по сравнению с чрескожным подходом привел к улучшению безрецидивной выживаемости и уменьшению локального рецидива опухоли [9].
В исследовании, сравнивающем открытый, лапароскопический и чрескожный доступы к опухолям печени, не было различий в смертности, серьезных осложнениях или общей выживаемости; но открытый по сравнению с чрескожным подходом привел к улучшению безрецидивной выживаемости и уменьшению локального рецидива опухоли [9]. Недостатками чрескожной методики являются отсутствие визуализации небольших поверхностных опухолей или более глубоких опухолей, которые лучше идентифицировать при открытой методике. Чрескожная РЧА показала отличные результаты при небольших новообразованиях < 3 см в печени, легких или почках. Однако при чрескожном доступе при больших опухолях [10] и опухолях, расположенных в непосредственной близости от крупных сосудов, таких как воротная вена, частота местных рецидивов выше.
Недостатками чрескожной методики являются отсутствие визуализации небольших поверхностных опухолей или более глубоких опухолей, которые лучше идентифицировать при открытой методике. Чрескожная РЧА показала отличные результаты при небольших новообразованиях < 3 см в печени, легких или почках. Однако при чрескожном доступе при больших опухолях [10] и опухолях, расположенных в непосредственной близости от крупных сосудов, таких как воротная вена, частота местных рецидивов выше.
Визуализация
 Однако эти гиперэхогенные области не полностью параллельны зоне абляции. Чтобы определить степень некроза после РЧА в странах за пределами США, во время процедуры используется контрастное УЗИ для проверки полной аблации и необходимости повторного лечения в данной ситуации [12]. В Соединенных Штатах обычно используется последующая КТ или МРТ с контрастным усилением, при этом успешно аблированные области не улучшаются. Тонкий контрастирующий ободок, представляющий либо воспаление, либо геморрагическую грануляционную ткань, может окружать зону абляции в течение нескольких недель после лечения [13].Последующее наблюдение может проводиться с помощью КТ, МРТ или позитронно-эмиссионной томографии, в зависимости от типа, размера и местоположения опухоли.
Однако эти гиперэхогенные области не полностью параллельны зоне абляции. Чтобы определить степень некроза после РЧА в странах за пределами США, во время процедуры используется контрастное УЗИ для проверки полной аблации и необходимости повторного лечения в данной ситуации [12]. В Соединенных Штатах обычно используется последующая КТ или МРТ с контрастным усилением, при этом успешно аблированные области не улучшаются. Тонкий контрастирующий ободок, представляющий либо воспаление, либо геморрагическую грануляционную ткань, может окружать зону абляции в течение нескольких недель после лечения [13].Последующее наблюдение может проводиться с помощью КТ, МРТ или позитронно-эмиссионной томографии, в зависимости от типа, размера и местоположения опухоли. Исключениями из правила отступа в 1 см могут быть такие органы, как почки, в которых сохранение нормальной почечной паренхимы было бы приоритетом, или когда целью лечения является уменьшение объема опухоли для паллиативного лечения или облегчения нейроэндокринных симптомов, или когда окружающие жизненно важные структуры ограничивают степень абляции.
Исключениями из правила отступа в 1 см могут быть такие органы, как почки, в которых сохранение нормальной почечной паренхимы было бы приоритетом, или когда целью лечения является уменьшение объема опухоли для паллиативного лечения или облегчения нейроэндокринных симптомов, или когда окружающие жизненно важные структуры ограничивают степень абляции. Осложнения
ОПЫТ АБЛАТИВАЦИИ ТВЕРДЫХ ОПУХОЛЕЙ
Печень
 В настоящее время РЧА считается методом лечения первой линии для локального контроля гепатоцеллюлярной карциномы у пациентов с циррозом по Чайлд-Пью B или более высокой степенью, когда резекция может иметь более высокую связанную с ней смертность. Он показан пациентам с 3 или менее опухолями размером 3 см или меньше (Миланские критерии) [18].Недавно было показано, что он превосходит чрескожную инъекцию этанола в отношении выживаемости и местного рецидива [19]. Эквивалентность хирургической резекции у пациентов, удовлетворяющих Миланским критериям, остается спорной. Проспективное рандомизированное исследование и большой ретроспективный анализ, сравнивающий локальные аблативные методы с хирургической резекцией у пациентов с небольшими солитарными опухолями, стадия Т1, не выявили различий в общей выживаемости между РЧА и хирургической резекцией [20-22]. Меньшие обсервационные исследования продемонстрировали аналогичные результаты [11].Метаанализ, сравнивающий РЧА с резекцией печени во всех подгруппах пациентов, показал улучшение общей 3- и 5-летней выживаемости и безрецидивной выживаемости, а также снижение местных рецидивов у пациентов, перенесших резекцию печени [23].
В настоящее время РЧА считается методом лечения первой линии для локального контроля гепатоцеллюлярной карциномы у пациентов с циррозом по Чайлд-Пью B или более высокой степенью, когда резекция может иметь более высокую связанную с ней смертность. Он показан пациентам с 3 или менее опухолями размером 3 см или меньше (Миланские критерии) [18].Недавно было показано, что он превосходит чрескожную инъекцию этанола в отношении выживаемости и местного рецидива [19]. Эквивалентность хирургической резекции у пациентов, удовлетворяющих Миланским критериям, остается спорной. Проспективное рандомизированное исследование и большой ретроспективный анализ, сравнивающий локальные аблативные методы с хирургической резекцией у пациентов с небольшими солитарными опухолями, стадия Т1, не выявили различий в общей выживаемости между РЧА и хирургической резекцией [20-22]. Меньшие обсервационные исследования продемонстрировали аналогичные результаты [11].Метаанализ, сравнивающий РЧА с резекцией печени во всех подгруппах пациентов, показал улучшение общей 3- и 5-летней выживаемости и безрецидивной выживаемости, а также снижение местных рецидивов у пациентов, перенесших резекцию печени [23]. Однако у пациентов с опухолями менее 3 см общая выживаемость была сопоставимой. Было показано, что у пациентов с большими опухолями (> 3 см) комбинация химиоэмболизации с РЧА превосходит только РЧА в улучшении выживаемости [24, 25]. Это основано на гипотезе о том, что РЧА приводит к зоне воспаления, которую затем можно стратегически использовать для адресной доставки химиотерапевтических агентов посредством химиоэмболизации .
Однако у пациентов с опухолями менее 3 см общая выживаемость была сопоставимой. Было показано, что у пациентов с большими опухолями (> 3 см) комбинация химиоэмболизации с РЧА превосходит только РЧА в улучшении выживаемости [24, 25]. Это основано на гипотезе о том, что РЧА приводит к зоне воспаления, которую затем можно стратегически использовать для адресной доставки химиотерапевтических агентов посредством химиоэмболизации . Рандомизированные контрольные исследования, непосредственно сравнивающие РЧА с резекцией печени при операбельном заболевании, еще предстоит провести.
Рандомизированные контрольные исследования, непосредственно сравнивающие РЧА с резекцией печени при операбельном заболевании, еще предстоит провести. Поэтому очевидно, что хирургическая резекция остается золотым стандартом; но для тех, кто не является кандидатом на операцию, ценна такая альтернатива, как РЧА.
Поэтому очевидно, что хирургическая резекция остается золотым стандартом; но для тех, кто не является кандидатом на операцию, ценна такая альтернатива, как РЧА. Таблица 2
Ref. пациентов (опухоли) N срединный размер опухоли (см) химиотерапия % полное облучение Местный рецидив Общая выживаемость 90 476 1 год 3 года 5 год Abdalla et al[26] 57 (110) для RFA 2.  5
5 № NR 0 NR 9% для RFA NR 37% для RFA43% для HR + RFA NR 90 для HR 5% для HR + RFA 93% для HR 93% 101 для RFA + HR 2% для HR Сиперштейн и др.  [27]
[27] 234 (665) 3.9 (значит) да 80% до RFA L NR NR 2 18% 2 Park et al [28] 30 для RFA 2.0 для RFA NO 73% после RFA P P NR 23% для RFA NR NR 19% 2 для RF1 59 для HR 3.  1 для HR
1 для HR 81% после HR 2% для HR 48% 2 для HR Abitabile et al [54] 47 (147) 2 2 после RFA O, P 97% 9% для общего 9% 1 57% 1 21% 1 9029 0% -5% для <3 см Gillams et al [55] 167 (167) 3.  9 (значит)
9 (значит) да 80% до RFA P NR % 99% 1 58% 1 30% 1 91% 2 2 28% 2 25% 2 Jakobs et al [56] 68 (183) 2.  28 (Среднее)
28 (Среднее) № № 78% параллельно или после P NR 18.00% 96% 2 71% 2 100 (507) 3.0 (среднее значение) NR O, L, P 7% 90% 42% 31% Schindera et al [58] 14 (20) 1.8 № 3 № NR P 89% 15% 72% 2 60% 2 NR White et et al [59] 30 (56) 3.  0 (0.8-7)
0 (0.8-7) № 36% до, 50% после P 89% 17% 75% 2 45% 2 NR Solbiati et al [60] 117 (179) 2.6 Да 72% Parallel P 98% 39% 93% 2 46% 2 NR Легкие
 Хирургическая резекция остается золотым стандартом радикального лечения первичного рака легкого и злокачественных метастазов. Однако только около 30% пациентов с первичным раком легкого подлежат хирургическому лечению на момент постановки диагноза из-за плохого функционального состояния и хронической обструктивной болезни легких [29]. У пациентов с легочными метастазами множественные поражения и поздняя стадия обычно исключают лечебную хирургическую резекцию.
Хирургическая резекция остается золотым стандартом радикального лечения первичного рака легкого и злокачественных метастазов. Однако только около 30% пациентов с первичным раком легкого подлежат хирургическому лечению на момент постановки диагноза из-за плохого функционального состояния и хронической обструктивной болезни легких [29]. У пациентов с легочными метастазами множественные поражения и поздняя стадия обычно исключают лечебную хирургическую резекцию.
 В большинстве исследований, в которых сравниваются исходы в зависимости от размера аблированной опухоли, у пациентов с опухолями размером менее 3 см были более длинные медианы интервалов без прогрессирования и общая выживаемость [35].
В большинстве исследований, в которых сравниваются исходы в зависимости от размера аблированной опухоли, у пациентов с опухолями размером менее 3 см были более длинные медианы интервалов без прогрессирования и общая выживаемость [35]. Таблица 3
Ref. Пациенты (опухоли) N Среднее размера опухоли (см) Median Местный прогресс Бесплатный интервал Общая выживание Осложнения 9000 2 Yr 3 YR Ambrogi et al [1] 54 (64) 2 .  4
4 40 для NSCLC <3 см - 15.8 MO 72% для NSCLC 1 46% для NSCLC 1 30% для NSCLC 1 6 для PTX 24 для Mets > 3 см — 6.6 MO 88% для MO 9 88% для MET 1 72% для Mets 1 NR для Mets 1 1 для грудной клетки Гематома Kim et al[30] 8 для RFA 3.  66 Для RFA
66 Для RFA Все этап I NSCLC NR 88% для RFA 50% для RFA 25% для RFA 1 для PTX 14 для SR 3,99 для SR 93% для SR 77% для SR 67% для SR 4 для HEMOPTYSIS 4 для HEMOPTYSIS 153 (189) 153 (189) 2,7 75 для сцены Я NSCLC <3 см - 45 MO 78% для NSCLC 57% для NSCLC 1 36% для NSCLC 1 18 для PTX 57 Для Mets > 3 см — 12 MO 70% для Met 54% для Mets 1 44% для Mets 1 5 для HEMOPTYSIS 4 для смерти Chua et al [37] 148 4 108 для CRCM 11 MO NR NR 60299 60% 66 для PTX Другое 16 для плеврального выпота 40 для Mets 1 для vleeding Lencioni et al[61] 106 (183) 3.  5
5 33 33 для NSCLC NR 70% для NSCLC 48% для NSCLC 27 для PTX 73 для Mets 93 для METS 99% для CRCM 66% для CRCM 4 для выпуска 92% для других 92% для других 64% для других Yan et al [62] 55 2.  1
1 Все CRCM NR 85% 64% 64% 16% 16% PTX / 9, требующие дренажа Для HEMOPTYSIS Hiraki et al [63] 20 2.4 Все этап I NSCLC 9 MO 90% 84% 74% 13 для PTX / 1, требующие дренажа  5% и 0% соответственно[36]. Большинство осложнений торакальной РЧА незначительны, наиболее часто встречающимися являются пневмоторакс и плевральный выпот (4,5%-61%) и кровохарканье. Другие включают боль, лихорадку и пневмонию. Несмотря на высокую заболеваемость пневмотораксом, лишь немногим, 11%, требуется плевральный дренаж [36]. Частота пневмоторакса увеличивается по мере увеличения количества удаленных поражений [37].
5% и 0% соответственно[36]. Большинство осложнений торакальной РЧА незначительны, наиболее часто встречающимися являются пневмоторакс и плевральный выпот (4,5%-61%) и кровохарканье. Другие включают боль, лихорадку и пневмонию. Несмотря на высокую заболеваемость пневмотораксом, лишь немногим, 11%, требуется плевральный дренаж [36]. Частота пневмоторакса увеличивается по мере увеличения количества удаленных поражений [37]. Грудь
 Ответ оценивали с помощью МРТ до и после процедуры, которая лучше коррелировала с патологическим ответом, чем УЗИ [38].Для оценки гистопатологического ответа использовали окрашивание HE, иммуногистохимию с CK 18/8 или анализ жизнеспособности клеток никотинамидадениндинуклеотид-диафоразой. Есть несколько исследований, в которых сообщалось, что окрашивание HE может быть недостаточным для оценки гистопатологического ответа, поскольку оно дает широкий спектр некрозов, и что методы, которые оценивают жизнеспособность клеток, лучше [38,39]. Полный коагуляционный некроз был достигнут у 80-100% пациентов, при этом ожог кожи был наиболее частым осложнением в очень небольшой группе пациентов.
Ответ оценивали с помощью МРТ до и после процедуры, которая лучше коррелировала с патологическим ответом, чем УЗИ [38].Для оценки гистопатологического ответа использовали окрашивание HE, иммуногистохимию с CK 18/8 или анализ жизнеспособности клеток никотинамидадениндинуклеотид-диафоразой. Есть несколько исследований, в которых сообщалось, что окрашивание HE может быть недостаточным для оценки гистопатологического ответа, поскольку оно дает широкий спектр некрозов, и что методы, которые оценивают жизнеспособность клеток, лучше [38,39]. Полный коагуляционный некроз был достигнут у 80-100% пациентов, при этом ожог кожи был наиболее частым осложнением в очень небольшой группе пациентов. Таблица 4
Ref. пациентов диапазон опухоли (см) средний размер опухоли (см) полная коагуляция некроз N (%) резекция оценка клеток жизнеспособность Осложнения Burak et al[38] 10 0.  8-1.6
8-1.6 1.2 9 ( 9 (90) задержка он CK8 / 18 NORE Singletary et al [40] 29 ≤ 2.0 — 25 (86) Немедленный Он Nadh-Diphorase 1 Skin Burn 52 0.52.0 1.3 52 (100) Задержка NR 1 ожог кожи Khatri et al[64] 15 0.  8-1.5
8-1.5 1.28 1,28 13 (93) Немедленный он Nadh-Diphorase 2 Skin Pucking Noguchi et al [65] 10 0.5-2,0 1.1 10 (100) Немедленный он Nadh-diphorase NORE 20 0,6-2,0 0,6-2,0 1.2 21 (100) Немедленно Он нада диафораза Нет Hayashi et al[67] 22 0.  5-2.6
5-2.6 0.9 (Median) 19 (86) задержка он нада 1 Skin Burn 26 26 0,7-3,0 1.8 25 (96) Неотложная ГЭ НАДН-диафораза 1 ожог кожи  Кроме того, статус рецепторов эстрогена и прогестерона, ее статус 2, степень, гистология и потребность в химиотерапии должны были быть известны до РЧА, поскольку в случае 100% успеха после процедуры не будет остаточных опухолевых клеток. Поверхностные опухоли в пределах 1 см от кожи также являются относительным противопоказанием из-за повышенного риска ожогов кожи. В исследованиях использовались различные стратегии для минимизации ожогов кожи, включая охлаждение груди стерильными пакетами со льдом и подкожные инъекции стерильного физиологического раствора или раствора с высоким сопротивлением для удаления опухоли с кожи.Кроме того, предоперационная химиотерапия является противопоказанием, поскольку она может привести к недооценке размера опухоли и оставить скрытые очаги резидуальной карциномы [40].
Кроме того, статус рецепторов эстрогена и прогестерона, ее статус 2, степень, гистология и потребность в химиотерапии должны были быть известны до РЧА, поскольку в случае 100% успеха после процедуры не будет остаточных опухолевых клеток. Поверхностные опухоли в пределах 1 см от кожи также являются относительным противопоказанием из-за повышенного риска ожогов кожи. В исследованиях использовались различные стратегии для минимизации ожогов кожи, включая охлаждение груди стерильными пакетами со льдом и подкожные инъекции стерильного физиологического раствора или раствора с высоким сопротивлением для удаления опухоли с кожи.Кроме того, предоперационная химиотерапия является противопоказанием, поскольку она может привести к недооценке размера опухоли и оставить скрытые очаги резидуальной карциномы [40]. Очень немногие исследования оценивали РЧА как альтернативу хирургической резекции. Oura и коллеги [41] сообщили о своем опыте лечения 52 пациентов со средним размером опухоли 1.3 см (диапазон 0,5-2,0 см), с РЧА после биопсии сторожевого лимфатического узла. Не было местно-регионарного или отдаленного рецидива после среднего наблюдения в течение 15 месяцев (диапазон 6-30 месяцев).
Очень немногие исследования оценивали РЧА как альтернативу хирургической резекции. Oura и коллеги [41] сообщили о своем опыте лечения 52 пациентов со средним размером опухоли 1.3 см (диапазон 0,5-2,0 см), с РЧА после биопсии сторожевого лимфатического узла. Не было местно-регионарного или отдаленного рецидива после среднего наблюдения в течение 15 месяцев (диапазон 6-30 месяцев). Развитие технологий обработки изображений будет способствовать такому прогрессу. Тем не менее, поскольку все больше случаев рака молочной железы диагностируется при меньшем размере, сфокусированная абляция под визуальным контролем может свести к минимуму разрушение нормальной ткани молочной железы и, таким образом, может положительно повлиять на косметический эффект.
Развитие технологий обработки изображений будет способствовать такому прогрессу. Тем не менее, поскольку все больше случаев рака молочной железы диагностируется при меньшем размере, сфокусированная абляция под визуальным контролем может свести к минимуму разрушение нормальной ткани молочной железы и, таким образом, может положительно повлиять на косметический эффект. Почка
Противопоказания включают ожидаемую продолжительность жизни менее одного года, наличие отдаленных метастазов, опухоли > 5 см или опухоли в воротах или центральной собирательной системе. Исследования стабильно показывают 91–97% успеха первой аблации небольших (< 3–4 см) экзофитных периферически расположенных опухолей (таблица). Это связано с тем, что периферически расположенные опухоли окружены периренальным жиром, который обеспечивает изоляцию и позволяет достичь высоких температур, необходимых для успешной аблации.И наоборот, внутригрудной кровоток создает эффект теплоотвода, что затрудняет лечение центральных опухолей. Безрецидивная выживаемость варьирует от 79% до 91% при подтвержденном биопсией почечно-клеточном раке, в то время как 3- и 5-летняя выживаемость рака колеблется от 95% до 100% в нескольких долгосрочных исследованиях. Таблица 5
Ref. 
пациентов (опухоли) n среднего размера опухоли (см) RCC RCC Recurnence Free выживание Общая выживание ( YR) Рак конкретного выживания (YR) 1 3 5 3 5 Трейси и др.  [69]
[69] 208 (243) 9 9 298 79% 99% 97% 90% на 3 года 2 99% 1 93% 1 85% 95% для RCC 99% для RCC NR Levinson et al [70] 3 31 (34) P, L P, L 2.1 58% 91% 80% на 5 YR 2 NR NR 63% для всех 3 58% для ПКР 4 NR 100% для всех 100% для ПКР 4 для паранефральной гематомы; 1 при ожоге печени; 1 для смерти от пневмонии Zagoria et al[71] 41 (48) P 2.  6
6 100% NR 88% на 5 YR NR NR NR 66% NR NR 2 для пневмоторакса Нет дренажа; 2 для уректорных стриктур STRENT et al [72] 40 P, L P, L 2.4 81% 97% 91% на 3 года 2 NR NR NR 100% для RCC NR 2 для несовершеннолетних; 3 для крупных Опухоли костей и метастатические поражения костей
 Выполняется у пациентов с типичными клиническими и рентгенологическими характеристиками остеоид-остеомы (рентгенопрозрачный очаг, окруженный реактивным склерозом) для лечения болей в костях. Первоначально он успешен у 73-98% пациентов с 92-100% вторичным успехом, и у большинства пациентов наблюдается облегчение боли в течение первых 1-2 недель лечения [46-49]. Частота осложнений минимальна, наиболее распространены некроз кожи и ожоги. Было продемонстрировано, что она сопоставима с хирургической резекцией в отношении рецидивов [50].РЧА также была описана в сообщениях о случаях лечения других доброкачественных опухолей костей.
Выполняется у пациентов с типичными клиническими и рентгенологическими характеристиками остеоид-остеомы (рентгенопрозрачный очаг, окруженный реактивным склерозом) для лечения болей в костях. Первоначально он успешен у 73-98% пациентов с 92-100% вторичным успехом, и у большинства пациентов наблюдается облегчение боли в течение первых 1-2 недель лечения [46-49]. Частота осложнений минимальна, наиболее распространены некроз кожи и ожоги. Было продемонстрировано, что она сопоставима с хирургической резекцией в отношении рецидивов [50].РЧА также была описана в сообщениях о случаях лечения других доброкачественных опухолей костей. От 90 до 95 процентов пациентов, получавших РЧА, испытывают клинически значимое уменьшение боли, которое можно наблюдать в течение первой недели лечения и которое длится до 24 недель [52, 53]. Частота осложнений минимальна и может варьироваться от кровотечения, патологических переломов, ожогов кожи и мышц и повреждения соседних нервно-сосудистых структур [46].
От 90 до 95 процентов пациентов, получавших РЧА, испытывают клинически значимое уменьшение боли, которое можно наблюдать в течение первой недели лечения и которое длится до 24 недель [52, 53]. Частота осложнений минимальна и может варьироваться от кровотечения, патологических переломов, ожогов кожи и мышц и повреждения соседних нервно-сосудистых структур [46]. Глобальное влияние карантина из-за COVID-19 на загрязнение воздуха в городах | Elementa: Наука антропоцена
Разбавление Исправлено East Asia 26 января 26 февраля 2016-2020 гг. № 2 , SO 2 , CO и PM 2.5 PBLH (SU et al., 2020) (2020) Разбавление исправлено Северная Америка 1 января 30 апреля 2019-2020 гг. № 2 SZA, WS и WD (Goldberg et al., 2020) Разбавление исправлено с CO East Asia 1 января 14 марта 2020 NR-PM 1 — (Xu et al .  , 2020a)
, 2020a) Отношения трассер-трассер Восточная Азия 1 января–31 марта 2012–2019 PM 2.5 — — (Sun et al., 2020) Benchmarking Северная Америка Северная Америка 14 апреля 9 апреля 2019-2020 гг. CO, NO 2 и PM 2.5 Т и осад. (Tanzer-Gruener et al., 2020) Seneasalize Северная Америка 1 января 1 января 27 апреля 2015-2020 гг. PM 2.5 , № 2 , № x , а O 3 — — — (Adams, 2020) Desicalize East Asia 1 января 31 мая 2005-202099999 № 2 и AOD — (Diamond and Wood, 2020) desessalize Южная Азия и Восточная Азия 1 апреля 1 апреля 2016-2019 гг. 
2016-2019 № 2 , SO 2 , и CO — (Метя и др., 2020) Индексы дисперсии Восточная Азия 26 января–25 февраля 2013–2020 PM. 2.5 , PM 10 , так что 2 , CO, NO 2 , и O 3 WS, сдвиг ветра, потенциал T, и RH (Ван и Чжан, 2020) Обратная траектория Восточная Азия 1 января–26 февраля 2019–2020 PM 2.5 HYSPLIT, 2020) Back-Trajectory и PMF East Asia 12 января 12 января — 2 20199 вечера 2.5 GDAS (Cui et al., 2020) -Транжек Южная Америка 1 апреля 1 апреля 9 апреля 2020 CO, № 2 , O 3 , VOC и PM 10 Hysplit (Siciliano et al.  , 2020b)
, 2020b) Обратный траекторный и кластерный анализ Восточная Азия 23 января–8 апреля 2020 PM 2.5 , так 2 , № 2 , CO, и O 3 Hysplit (Zhao et al., 2020A) Анализ обратной траектории East Asia 29 февраля 2000-20209 Aod Hysplit Hysplit (Shen et al., 2021) Машина обучения и PMF East Asia 23 января 22 февраля 2019 вечера 2,5 T, P, WS, RH, PBLH и радиация (Zheng et al., 2020) Нормализованный дисперсионный PMF East Asia East Asia 1 февраля 1 февраля 9 2020 г. PM 2,5 PM T T, WS, PBLE и радиация (Dai et al.  , 2020 )
, 2020 ) Анализ кластера Южная Азия Южная Азия 25 марта 15 мая 2017-202099998 , № 2 , SO 2 , O 3 , PM 10 и PM 2,5 T (Bera et al., 2020) Многофакторная регрессия East Asia East Asia января 23 марта 2019-2020 гг. 2019-2020 гг. SO 2 , PM 2.5 , PM 10 , № 2 , и CO WS, дождь и снег (Bao and Zhang, 2020) Метувариатская регрессия Северная Америка 25 марта 2 марта 2017-2020 гг. PM 2.5 , № 2 , и O 3 WS, T и осад. (Jia et al., 2020A) Метувариатская регрессия Европа 1 января 27 марта 1 марта 2017-2020 гг. № 2 и PM 10 T, WS и Precip . 
(CameLetti, 2020) Метувариатская регрессия Южная Америка, Северная Америка, и Европа 1 марта 31 марта 2015-2020 гг. PM 2.5 , CO, NO 2 , а также O 3 T, RH, WS и осад. . , T, RH, точка росы, стабильность, WS и осадки. (Lei et al., 2020) Метувариатская регрессия Global 1 января 15 мая 15 мая 2013 г. № № 2 , PM 2.5 , а o 3 T, Rh, Precip., И WS (Venter et al., 2020) Метувариатская регрессия Северная Америка 17 февраля — 31 мая 31 мая 20 мая BC, PM 2.5 , NO, NO 2 , NO x , CO и UFP T, RH, осадки, WS и WD 9020 2 920 et 8 al)8 al)8 et 44 (Xiang) Машина Обучение Европа Европа 1 января 23 апреля 2010-20999999949 2013-2020209999 № 2 T2, WS, U10, V10, P, Облачное покрытие, Излучение, УФ и PBLH Пететин и др.  , 2020)
, 2020) Машина Обучение East Asia 1 апреля 1 апреля 26 апреля 2020 г. № 2 , PM 2.5 и O 3 WS, WD, T, RH, и p (Wang et al., 2020e) Машина обучения Европа 1 марта 31 мая 31 марта 2015-2019 201544 № 2 , O 3 , PM 10 , и PM 2,5 WS, WD, P, RH, T и радиация (Wyche et al., 2020) Разница в разных методах Global 1 июля 1 июля 70298 2020199999 № 2 , PM 10 , SO 2 , PM 2.5 , CO, и O 3 T, WS, и RH (Liu et al., 2021A) Разница в разницах Южная Азия 25 марта 9 мая 2019-202099998 гг. PM 2,5 , PM 10 , NO 2 , CO и SO 2 T, WS и RH (Навинья и др.  , 2020)
, 2020) East Method East Asia East Asia 1 марта 2019-2020 гг. 2019-2020 гг. Индекс качества воздуха, PM 2.5 , CO, NO 2 , PM 10 , так что 2 , и O 3 T, Precip., И снег (он et al., 2020) Генерализованная добавка модель Европа 15 апреля 30298 2015–2019 НО 2 и О 3 Т2, У10, В10, Z500, удельная влажность, радиация, осадки. (Ordóñez et al., 2020) Генерализованная добавка модели Европа 10 марта 2015-20199 2015-2019 9198 Нет, № 2 , № x , O 3 , PM 10 , и PM 2 , и PM 2.5 WS, WD, и T (ROPKINS и TATE, 2020) Сердечная недостаточность с сохраненной фракцией выброса: диагностика и управление
 Borlaug BA ,
Паулюс В.Дж.Сердечная недостаточность с сохраненной фракцией выброса: патофизиология, диагностика, лечение. Европейское сердце J .
2011;32(6):670–679….
Borlaug BA ,
Паулюс В.Дж.Сердечная недостаточность с сохраненной фракцией выброса: патофизиология, диагностика, лечение. Европейское сердце J .
2011;32(6):670–679…. Тираж .
2009;119(24):3070–3077.
Тираж .
2009;119(24):3070–3077. Оценка медицинских технологий .
2009;13(32):1–207.
Оценка медицинских технологий .
2009;13(32):1–207. Открытие богатого генофонда коронавирусов, связанных с атипичной пневмонией летучих мышей, позволяет по-новому взглянуть на происхождение коронавируса атипичной пневмонии (2017) Обнаружение богатого генофонда коронавирусов, связанных с атипичной пневмонией летучих мышей, дает новое представление о происхождении коронавируса атипичной пневмонии. ПЛОС Патог 13(11):
е1006698.
https://doi.org/10.1371/журнал.ppat.1006698
Редактор: Кристиан Дростен, Charite Universitatsmedizin Berlin, GERMANY
Получено: 10 февраля 2017 г.; Принято: 17 октября 2017 г .; Опубликовано: 30 ноября 2017 г.
Авторское право: © 2017 Hu et al. Это статья с открытым доступом, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Доступность данных: Все соответствующие данные содержатся в документе и в его файлах вспомогательной информации. Полные геномные последовательности 11 коронавирусов, связанных с SARS летучих мышей, недавно идентифицированных в этом исследовании, были депонированы в базе данных GenBank, и им были присвоены номера доступа от KY417142 до KY417152 соответственно.
Финансирование: Эта работа была совместно профинансирована Национальным фондом естественных наук Китая (812, 31621061) для ZLS, Китайского мегапроекта по инфекционным заболеваниям (2014ZX10004001-003) для ZLS, Специального проекта научной и технологической базы (2013FY113500) в YZZ и ZLS от Министерства науки и технологий Китая, Стратегической приоритетной исследовательской программы Китайской академии наук (XDPB0301) в ZLS, Национальных институтов здравоохранения (NIAID R01AI110964), USAID Emerging Pandemic Threats (EPT) PREDICT программа для PD и ZLS, программа CAS Pioneer Hundred Talents для JC, грант NRF-CRP (NRF-CRP10-2012-05) для LFW и WIV Стратегическая программа «Один-три-пять» (WIV-135-TP1) для JC и ЗЛС.Спонсоры не участвовали в разработке исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявили об отсутствии конкурирующих интересов.
Введение
Тяжелый острый респираторный синдром (ТОРС) — это тяжелое эмерджентное вирусное заболевание с высокой летальностью, характеризующееся лихорадкой, головной болью и тяжелыми респираторными симптомами, включая кашель, одышку и пневмонию [1]. Из-за его высокой контагиозности среди людей после его первого появления на юге Китая в конце 2002 г. он быстро привел к глобальной пандемии в 2003 г. и был отмечен как одна из наиболее значительных угроз общественному здравоохранению в 219009-м -м веке [2, 3].Возбудитель, коронавирус SARS (SARS-CoV), ранее относился к группе 2b CoV и в настоящее время является представителем линии B рода Betacoronavirus семейства Coronaviridae [4]. Он имеет аналогичную организацию генома с другими коронавирусами, но обладает уникальной геномной структурой, которая включает ряд специфических дополнительных генов, включая ORF3a, 3b, ORF6, ORF7a, 7b, ORF8a, 8b и 9b [5,6].
Изначально предполагалось, чтопальмовых циветты в маске ( Paguma larvata ) являются источником животного происхождения SARS-CoV [7,8].Однако, поскольку большое количество генетически разнообразных коронавирусов, связанных с атипичной пневмонией (SARSr-CoV), было обнаружено у нескольких видов подковоносов (род Rhinolophus ) из разных районов Китая и Европы после атипичной пневмонии, это преимущественно считается что SARS-CoV возник у подковоносых летучих мышей, а циветты выступали в качестве промежуточного амплифицирующего и передающего хозяина [9–16]. Недавно мы сообщили о четырех новых SARSr-CoV от китайских подковоносых летучих мышей, которые имели гораздо большее сходство геномной последовательности с эпидемическими штаммами, особенно в их S-гене, из которых два штамма (названные WIV1 и WIV16) были успешно культивированы 90 265 in vitro 90 266 [90 266]. 17,18].Было продемонстрировано, что эти недавно идентифицированные SARSr-CoV используют тот же клеточный рецептор (ангиотензинпревращающий фермент-2 [ACE-2]), что и SARS-CoV, и эффективно реплицируются в первичных клетках дыхательных путей человека [17–19].
Несмотря на совокупные данные о появлении SARS-CoV у летучих мышей, все описанные до сих пор SARSr-CoV летучих мышей явно отличаются от SARS-CoV геном S и/или одним или несколькими дополнительными генами, такими как ORF3 и ORF8, что позволяет предположить, что они вероятно, не являются прямым предком SARS-CoV.Таким образом, остается критический пробел в нашем понимании того, как и где SARS-CoV возник из резервуаров летучих мышей. Ранее мы сообщали о ряде SARSr-CoV летучих мышей с различными последовательностями белков S из одной пещеры в провинции Юньнань, включая четыре упомянутых выше штамма, наиболее тесно связанных с SARS-CoV [17,18]. Здесь мы сообщаем о последних результатах нашего 5-летнего продольного наблюдения за SARSr-CoV летучих мышей в этом единственном месте и систематического эволюционного анализа с использованием полноразмерных геномных последовательностей 15 штаммов SARSr-CoV (11 новых и 4 из предыдущих исследований).Эффективность использования человеком ACE2 и функции дополнительных генов ORF8 и 8a также оценивали для некоторых из вновь идентифицированных штаммов.
Результаты
Продолжающаяся циркуляция различных SARSr-CoV у летучих мышей из одного места
Мы провели пятилетнее лонгитудинальное наблюдение (с апреля 2011 г. по октябрь 2015 г.) за SARSr-CoV у летучих мышей из единственного места обитания вблизи города Куньмин, провинция Юньнань, Китай, где в основном жили подковоносые летучие мыши. В общей сложности было собрано 602 образца пищи (анальные мазки или фекалии), которые были протестированы на наличие CoV с помощью ОТ-ПЦР Pan-CoV, направленной на 440-нуклеотидный фрагмент RdRp, который является консервативным среди всех известных α- и β-CoV [20]. ].В общей сложности 84 образца дали положительный результат на CoV. Секвенирование ампликонов ПЦР выявило наличие SARSr-CoV в большинстве (64/84) CoV-положительных образцов (таблица 1). Идентификация видов-хозяев путем амплификации гена Cytb или ND1 позволила предположить, что большинство (57/64) положительных на SARSr-CoV образцов были получены от Rhinolophus sinicus , а остальные 7 образцов были получены от Rhinolophus ferrumequinum , . Rhinolophus affinis и из Aselliscus stoliczkanus , который принадлежит к семейству Hippoideridae .
На основании предварительного анализа частичных последовательностей RdRp все 64 последовательности SARSr-CoV летучих мышей показали высокое сходство между собой и с другими зарегистрированными SARSr-CoV и SARS-CoV летучих мышей от людей и циветт. Чтобы понять генетическое разнообразие этих SARSr-CoV летучих мышей, была амплифицирована и секвенирована наиболее вариабельная область гена S SARSr-CoV, соответствующая рецептор-связывающему домену (RBD) SARS-CoV. Из-за низкой вирусной нагрузки в некоторых образцах последовательности RBD были успешно амплифицированы только из 49 образцов.Эти последовательности RBD демонстрируют высокое генетическое разнообразие и могут быть разделены на две большие клады, каждая из которых включает несколько генотипов. Штаммы клады 1 имели одинаковый размер и более высокую идентичность аминокислотной (аа) последовательности с RBD SARS-CoV, в то время как клада 2 имела более короткий размер, чем S SARS-CoV, из-за двух делеций (5 и 12–13 аа соответственно) (S1 Инжир). Коинфекции двумя штаммами разных кладов были обнаружены в двух образцах, Rs3262 и Rs4087 (S1 Fig).
Геномная характеристика новых SARSr-CoV
На основании разнообразия последовательностей RBD были отобраны 11 новых штаммов SARSr-CoV, названных по аббревиатуре видов летучих мышей и идентификатору образца (Rs4081, Rs4084, Rs4231, Rs4237, Rs4247, Rs4255, Rs4874, Rs7327, Rs9401, Rf4092 и As6526). полноразмерное геномное секвенирование на основе количества образцов, генотипа RBD, а также времени отбора образцов.Для каждого генотипа RBD и при каждом отборе образцов отбирали по крайней мере один репрезентативный штамм. Размер генома этих новых SARSr-CoV варьировался от 29694 до 30291 нуклеотидов (нт). Это дало в общей сложности 15 полноразмерных геномов SARSr-CoV летучих мышей из этого единственного места (13 из R . sinicus и по одному из R . ferrumequinum и A . stoliczkanus 9026zkanus). , включая наши ранее зарегистрированные штаммы Rs3367, RsSHC014, WIV1 и WIV16 [17,18].Геномы всех 15 SARSr-CoV, циркулирующих в этой единственной пещере, имели 92,0–99,9% идентичности последовательностей нуклеотидов. Общая идентичность последовательности нуклеотидов между этими SARSr-CoV и SARS-CoV человека и циветты составляет от 93,2% до 96%, что значительно выше, чем наблюдаемая для SARSr-CoV летучих мышей, о которых сообщалось из других мест в Китае (88–93%) [9,10]. ,12,14,21,22]. Сходство последовательностей геномов среди 15 штаммов SARSr-CoV и SARS-CoV SZ3 было исследовано с помощью анализа Simplot (рис. 1). 15 SARSr-CoV отличаются высокой консервативностью и одинаково высоким сходством последовательностей с SARS-CoV в неструктурном гене ORF1a (96.идентичность последовательности нуклеотида от 6% до 97,1%, идентичность последовательности аа от 98,0% до 98,3%) и ORF1b (идентичность последовательности нуклеотида от 96,1% до 96,6%, идентичность последовательности аа от 99,0% до 99,4%). Напротив, значительное генетическое разнообразие показано в гене S (соответствует положениям генома SZ3 с 21477 по 25244) и ORF8 (соответствует положениям генома SZ3 с 27764 по 28132) (рис. 1).
Рис. 1. График сходства, основанный на полноразмерной последовательности генома циветты SARS CoV SZ3.
Полноразмерные геномные последовательности всех SARSr-CoV, обнаруженных у летучих мышей из пещеры, исследованной в этом исследовании, использовались в качестве эталонных последовательностей.Анализ был выполнен с использованием модели Кимуры, размера окна 1500 пар оснований и размера шага 150 пар оснований.
https://doi.org/10.1371/journal.ppat.1006698.g001
11 новых SARSr-CoV, идентифицированных в этом единственном месте, в целом имели схожую организацию генома с SARS-CoV и другими SARSr-CoV летучих мышей. В нашем предыдущем исследовании мы идентифицировали дополнительную ORF, называемую ORFx, присутствующую между ORF6 и ORF7 в штаммах WIV1 и WIV16 [18,23]. В этом исследовании ORFx также была обнаружена в геномах Rs7327 и Rs4874.По сравнению с таковой у WIV1 и WIV16 длина ORFx у Rs7327 и Rs4874 была увеличена до 510 нуклеотидов из-за делеции 2 нуклеотидов в поли-Т-последовательности, что привело к смещению рамки считывания (рис. 2 и рис. S2).
Рис. 2. Схематическая диаграмма, иллюстрирующая области генома или ORF с наибольшим разнообразием между различными изолятами SARS-CoV и SARSr-CoV.
Кодирующие области N-концевого домена (NTD) и рецептор-связывающего домена (RBD) шиповидного белка, ORF3a/b и ORF8 (8a/b) в геномах SARSr-CoV летучих мышей, очень похожих на таковые в геноме SARS-CoV обозначены черными прямоугольниками или стрелками, в то время как пустые прямоугольники или стрелки представляют соответствующие области с меньшим сходством последовательностей с последовательностями SARS-CoV.Делеции в RBD некоторых SARSr-CoV обозначены двумя вертикальными линиями.
https://doi.org/10.1371/journal.ppat.1006698.g002
Совместная циркуляция различных SARSr-CoV летучих мышей с последовательностями S, ORF8 и ORF3, аналогичными последовательностям SARS-CoV, в одном месте
Основное различие между SARS-CoV и большинством SARSr-CoV летучих мышей заключается в гене S. Белок S функционально разделен на две субъединицы, обозначенные как S1 и S2, которые отвечают за связывание с рецептором и слияние с клеточной мембраной соответственно.S1 состоит из двух доменов: N-концевого домена (NTD) и С-концевого домена (CTD), который также известен как RBD в SARS-CoV [24]. SARS-CoV и SARSr-CoV летучих мышей имеют высокую идентичность последовательностей в области S2, в отличие от области S1. Среди 15 SARSr-CoV, идентифицированных у летучих мышей в обследованной пещере, шесть штаммов с делециями в областях RBD (Rs4081, Rs4237, Rs4247, Rs4255, Rf4092 и As6526) показали 78,2–80,2% идентичности последовательностей с SARS-CoV в Белок S, в то время как остальные девять штаммов без делеций были гораздо более тесно связаны с SARS-CoV, с 90.от 0% (Rs4084) до 97,2% (Rs4874) идентичности последовательностей. Эти девять SARSr-CoV можно дополнительно разделить на четыре генотипа в соответствии с их последовательностями S1 (рис. 2): RsSHC014/Rs4084 показали больше генетических отличий от SARS-CoV как в области NTD, так и в области RBD; Последовательности RBD вирусов SARSr-CoV Rs7327, Rs9401 и WIV1/Rs3367, о которых сообщалось ранее, очень напоминали последовательности SARS-CoV. Однако они отличались от SARS-CoV, но сходны с RsSHC014 при ДНТ. Напротив, мы обнаружили новый SARSr-CoV, названный Rs4231, который имеет очень похожую последовательность NTD, но не RBD с SARS-CoV (рис. 2 и 3).Его S-белок показал 94,6-95% идентичность последовательности с последовательностями SARS-CoV человека и циветты (таблица S1). В этой пещере также присутствовали штаммы с NTD и RBD, высоко гомологичные штаммам SARS-CoV. В дополнение к WIV16, который мы описали ранее [18], также было обнаружено, что Rs4874 имеет белок S, наиболее близкий к S SARS-CoV (> 97% идентичности последовательности) из всех зарегистрированных на сегодняшний день SARSr-CoV летучих мышей (рис. 2 и 3). ). В дополнение к SARSr-CoV, подвергнутым полноразмерному секвенированию генома, мы также получили последовательности RBD и NTD из других образцов, собранных в этой пещере.Последовательности с высокой идентичностью RBD SARS-CoV были амплифицированы еще из 10 R . синикусов образцов. SARSr-CoV с данным генотипом RBD выявляли в разные сезоны на протяжении пяти лет. Штаммы, содержащие NTD, похожие на SARS-CoV, были обнаружены только в 2013 г. (таблица S2).
Рис. 3. Сравнение аминокислотной последовательности субъединицы S1 (соответствует а.о. 1–660 шиповидного белка SARS-CoV).
Домен связывания рецептора (аминокислоты 318–510) SARS-CoV и гомологичный регион SARSr-CoV летучих мышей обозначены красной рамкой.Ключевые аминокислотные остатки, участвующие во взаимодействии с человеческим ACE2, пронумерованы над выровненными последовательностями. SARS-CoV GZ02, BJ01 и Tor2 были выделены у пациентов на ранней, средней и поздней фазах вспышки атипичной пневмонии в 2003 г., соответственно. SARS-CoV SZ3 был идентифицирован у цивет в 2003 г. от р . sinicus собраны в Гуйчжоу и провинции Юньнань соответственно. SARSr-CoV Rf1 и JL2012 были идентифицированы из R . ferrumequinum собраны в провинциях Хубэй и Цзилинь соответственно. WIV1, WIV16, RsSHC014, Rs4081, Rs4084, Rs4231, Rs4237, Rs4247, Rs7327 и Rs4874 были идентифицированы из R . sinicus и Rf4092 из R . ferrumequinum в пещере, обследованной в данном исследовании.
https://doi.org/10.1371/journal.ppat.1006698.g003
ORF8 — еще один высоковариабельный ген среди различных штаммов SARS-CoV и SARSr-CoV [25,26].Мы сопоставили последовательности нуклеотидов ORF8 репрезентативных SARSr-CoV, обнаруженных в ходе этого наблюдения, с последовательностями других SARSr-CoV и SARS-CoV (рис. 4). Хотя WIV16, WIV1, Rs4231 и RsSHC014 были генетически ближе к SARS-CoV в гене S, они содержали одну ORF8 из 366 нуклеотидов без 29-нуклеотидной делеции, присутствующей в большинстве SARS-CoV человека, и имели последовательность только от 47,1% до 51,0% нуклеотидов. идентичность к SARS-CoV человека и циветты. Однако ORF8 штамма Rf4092 из R . ferrumequinum показал высокое сходство с циветтой SARS-CoV.Он обладал единственной длинной ORF8 той же длины (369 нт), что и у циветты SARS-CoV, штамм SZ3, с обнаруженными только 10 нуклеотидными мутациями и 3 аа-мутациями (рис. 4). Подобные последовательности ORF8 также были амплифицированы из других 7 образцов, собранных в пещере в период с 2011 по 2013 год, из обоих R . феррумхин и R . sinicus (таблица S2). ORF8 Rs4084 была очень похожа на Rf4092, но была разделена на две перекрывающиеся ORF8a и ORF8b из-за короткой 5-нуклеотидной делеции (рис. 2 и 4).Положение старт-кодонов и стоп-кодонов двух ORF соответствовало таковому в большинстве штаммов SARS-CoV человека. За исключением вставки из 8 а.о., штамм Rs4084 и SARS-CoV BJ01 демонстрировал идентичную последовательность а.о. ORF8a, и между их ORF8b наблюдались только три разных аминокислотных остатка (рис. 4). Насколько нам известно, Rs4084 был первым зарегистрированным SARSr-CoV летучих мышей, который напоминал поздние человеческие SARS-CoV как по организации гена ORF8, так и по последовательности.
Рис. 4. Выравнивание нуклеотидных последовательностей ORF8 или ORF8a/8b.
Старт-кодоны и стоп-кодоны ORF8, 8a и 8b отмечены черными прямоугольниками и прямой и обратной стрелками соответственно. Делеция, ответственная за расщепление ORF8a и 8b в SARS-CoV BJ01, Tor2 человека и SARSr-CoV Rs4084 летучей мыши, отмечена красными прямоугольниками. См. легенду к рис. 3, чтобы узнать о происхождении различных последовательностей, использованных в этом выравнивании.
https://doi.org/10.1371/journal.ppat.1006698.g004
Другим ключевым различием между геномами SARS-CoV и SARSr-CoV летучих мышей является кодирующая область ORF3 [10,17,21].Мы проанализировали последовательности ORF3a, амплифицированные из 42 образцов, и обнаружили, что большинство SARSr-CoV, тесно связанных с SARS-CoV в гене S, имеют более высокое сходство последовательностей ORF3a (96,4–98,9% идентичности аа) с SARS-CoV (рис. S3 и Таблица S2). ORF3b SARS-CoV, имеющая большую часть своей кодирующей последовательности с ORF3a, кодирует белок из 154 а.о. WIV16 (S4 рис). В текущем исследовании мы впервые идентифицировали неусеченную ORF3b (Rs7327), которая поддерживала сигнал ядерной локализации на своем С-конце.Более того, он имел 98,1% идентичности аминокислотной последовательности со штаммом Tor2 SARS-CoV только с тремя заменами аминокислот (рис. S4). Таким образом, Rs7327 представляет собой SARSr-CoV летучих мышей, наиболее похожий на SARS-CoV в известной на сегодняшний день области ORF3.
Рекомбинационный анализ
Полноразмерные геномные последовательности всех 15 SARSr-CoV из обследованной пещеры были проверены на наличие признаков потенциальной рекомбинации. Как график подобия, так и анализ бутскана выявили частые события рекомбинации среди этих штаммов SARSr-CoV.Было высказано предположение, что WIV16, ближайший предшественник человеческого SARS-CoV, известный на сегодняшний день [18], вероятно, является рекомбинантным штаммом трех SARSr-CoV, населяемых летучими мышами в одной и той же пещере, а именно WIV1, Rs4231 и Rs4081, с сильным Значение P (<10 −30 ). Точки разрыва были идентифицированы в положениях генома нуклеотидов 18391, 22615 и 28160 (фиг. 5А). В области генома между нуклеотидами 22615 и 28160, которая содержит область, кодирующую RBD и субъединицу S2 белка S, WIV16 был очень похож на WIV1, обладая 99% идентичностью последовательности.Напротив, в области между нуклеотидами 18391 и 22615, которая покрывает часть ORF1b и область, кодирующую NTD гена S, WIV16 показал существенно более близкое родство с Rs4231. Между тем, последовательности ORF1ab выше нуклеотида 18391 WIV16 демонстрировали наивысшее генетическое сходство (99,8% идентичности последовательности нуклеотида) с последовательностями Rs4081.
Рис. 5. Обнаружение потенциальных событий рекомбинации с помощью графика подобия и анализа загрузки.
(A) Полноразмерная последовательность генома SARSr-CoV WIV16 использовалась в качестве запрашиваемой последовательности, а WIV1, Rs4231 и Rs4081 — в качестве эталонных последовательностей.(B) Полноразмерная последовательность генома SARS-CoV SZ3 использовалась в качестве запрашиваемой последовательности, а SARSr-CoV WIV16, Rf4092 и Rs4081 — в качестве эталонных последовательностей. Все анализы проводились с использованием модели Кимуры, размера окна 1500 пар оснований и размера шага 150 пар оснований. Генная карта последовательностей генома запроса используется для позиционирования точек останова.
https://doi.org/10.1371/journal.ppat.1006698.g005
Доказательства события рекомбинации были также обнаружены в геноме нового SARSr-CoV Rs4084, который имел уникальную организацию генома с расщепленными ORF8a и 8b.Предполагалось, что штамм RsSHC014, о котором сообщалось ранее, и недавно идентифицированный штамм Rf4092 являются основным и второстепенным родителем Rs4084 соответственно (значение P < 10 -80 ). Точка останова была расположена на nt 26796 (рис. S5). В области ниже точки разрыва, включая ORF8, Rs4084 продемонстрировал тесную генетическую связь с Rf4092, разделяя 98,9% идентичности последовательности нуклеотидов, в то время как он разделял самую высокую идентичность последовательности нуклеотидов (99,4%) с RsSHC014 в большей части своего генома выше точки разрыва.
Когда циветта SARS-CoV SZ3 использовалась в качестве запрашиваемой последовательности на графике сходства и анализе загрузки, также были обнаружены свидетельства событий рекомбинации (рис. 5B). В области между двумя точками разрыва в геномных позициях нуклеотидов 21161 и 27766, включая ген S, наблюдалось более близкое генетическое родство между SZ3 и WIV16. Однако от положения нуклеотида 27766 к 3′-концу его генома вместо этого наблюдалось заметно близкое генетическое родство между SZ3 и Rf4092. Кроме того, во всем неструктурном гене SZ3 имеет аналогичную высокую идентичность последовательностей с WIV16 и Rf4092.Это указывает на то, что SARS-CoV циветты, вероятно, был потомком рекомбинанта предшественников WIV16 и Rf4092, или что обнаруженные в этой пещере вирусы SARSr-CoV, такие как WIV16 или Rf4092, могли быть потомками SARS-CoV. происхождение.
Филогенетический анализ
Филогенетические деревья были построены с использованием последовательностей нуклеотидов генов неструктурных белков ORF1a и ORF1b. В отличие от высокого генетического разнообразия гена S, почти все SARSr-CoV из обследованной нами пещеры летучих мышей были тесно сгруппированы и показали более тесную филогенетическую связь с SARS-CoV, чем большинство известных в настоящее время SARSr-CoV летучих мышей, обнаруженных в других местах, за исключением YNLF_31C и 34C, о которых недавно сообщалось у больших подковоносов из другого места в Юньнани [22] (рис. 6).Филогения SARSr-CoV в ORF1a и ORF1b, по-видимому, связана с их географическим распространением, а не с видами-хозяевами. Независимо от разных видов летучих мышей-хозяев, SARS-CoV и SARSr-CoV, обнаруженные у летучих мышей из юго-западного Китая (Юньнань, Гуйчжоу и провинция Гуанси), сформировали одну кладу, в которой все штаммы SARSr-CoV, демонстрирующие более тесную связь с SARS-CoV, были из Юньнани. SARSr-CoV, обнаруженные в юго-восточных, центральных и северных провинциях, таких как Гонконг, Хубэй и Шэньси, сформировали другую кладу, которая была филогенетически отдалена от SARS-CoV человека и циветты (рис. 6 и рис. S6).
Рис. 6.
Филогенетические деревья на основе нуклеотидных последовательностей ORF1a (A) и ORF1b (B). Деревья были построены методом максимального правдоподобия с использованием модели LG со значениями начальной загрузки, определенными в 1000 повторах. Показаны только бутстрапы > 50%. Шкала баров представляет 0,03 (A) и 0,02 (B) замены на положение нуклеотида. Rs, Rhinolophus sinicus ; Rf, Rhinolophus ferremequinum ; Rm, Rhinolophus macrotis ; Ra, Rhinolophus affinis ; Rp, Rhinolophus pusillus ; As, Aselliscus stoliczkanus ; Cp, Chaerephon plicata .Жирным шрифтом выделены вирусы SARSr-CoV, обнаруженные у летучих мышей из единственной пещеры, исследованной в этом исследовании. Последовательности, обнаруженные на юго-западе Китая, обозначены красным цветом.
https://doi.org/10.1371/journal.ppat.1006698.g006
Спасение SARSr-CoV летучих мышей и эксперименты с инфекционностью вируса
В текущем исследовании мы успешно культивировали дополнительный новый вирус SARSr-CoV Rs4874 из одного образца фекалий с использованием оптимизированного протокола и клеток Vero E6 [17]. Его белок S разделяет 99.9% идентичность последовательности с последовательностью ранее выделенного WIV16, и она была идентична WIV16 в RBD. Используя метод обратной генетики, который мы ранее разработали для WIV1 [23], мы сконструировали группу клонов инфекционных бактериальных искусственных хромосом (ВАС) с основой WIV1 и вариантами генов S из 8 различных SARSr-CoV летучих мышей. Только инфекционные клоны для Rs4231 и Rs7327 приводили к цитопатическим эффектам в клетках Vero E6 после трансфекции (S7 Fig). Остальные шесть штаммов с делециями в области RBD, Rf4075, Rs4081, Rs4085, Rs4235, As6526 и Rp3 (рис. S1), спасти не удалось, поскольку не наблюдалось цитопатических эффектов, а репликация вируса не могла быть обнаружена с помощью иммунофлуоресцентного анализа в клетках Vero E6. (S7 рис.).Напротив, когда клетки Vero E6 были инфицированы соответственно двумя успешно восстановленными химерными SARSr-CoV, WIV1-Rs4231S и WIV1-Rs7327S, и вновь выделенным Rs4874, эффективная репликация вируса была обнаружена при всех инфекциях (рис. 7). Чтобы оценить, могут ли три новых SARSr-CoV использовать человеческий ACE2 в качестве рецептора проникновения в клетку, мы провели исследования вирусной инфекционности с использованием клеток HeLa с экспрессией человеческого ACE2 или без нее. Все вирусы эффективно реплицировались в клетках, экспрессирующих ACE2 человека.Результаты были дополнительно подтверждены количественным определением вирусной РНК с использованием ОТ-ПЦР в реальном времени (фиг. 8).
Рис. 7. Заражение клеток Vero E6 вирусом SARSr-CoV летучих мышей WIV1, Rs4874, WIV1-Rs4231S и WIV1-Rs7327S.
(A) Успешное инфицирование было подтверждено окрашиванием иммунофлуоресцентными антителами с использованием кроличьих антител против нуклеокапсидного белка Rp3 SARSr-CoV. Столбцы (слева направо) показывают окрашивание ядер (синий), репликацию вируса (красный), а также репликацию ядер и вируса (объединенные изображения двойного окрашивания).(B) Кривые роста в клетках Vero E6 с MOI 1,0 и 0,01.
https://doi.org/10.1371/journal.ppat.1006698.g007
Рис. 8.
Анализ использования рецепторов с помощью иммунофлуоресцентного анализа (A) и ПЦР в реальном времени (B). Вирусная инфекционность Rs4874, WIV1-Rs4231S и WIV1-Rs7327S определялась в клетках HeLa с экспрессией ACE2 человека и без нее. Экспрессию ACE2 определяли с помощью козьего антитела против человеческого ACE2, а затем конъюгированного с флуоресцеинизотиоцианатом (FITC) ослиного IgG против козла.Репликация вируса была обнаружена с помощью кроличьего антитела против нуклеокапсидного белка SARSr-CoV Rp3, а затем конъюгированного с цианином 3 (Cy3) мышиного антикроличьего IgG. Ядра окрашивали DAPI (49,6-диамидино-2-фенилиндол). В столбцах (слева направо) показано окрашивание ядер (синий), экспрессия ACE2 (зеленый), репликация вируса (красный) и объединенное тройное окрашивание. изображений соответственно.
https://doi.org/10.1371/journal.ppat.1006698.g008
Активация активирующего фактора транскрипции 6 (ATF6) белками ORF8 различных SARSr-CoV летучих мышей
Индукцию ATF6-зависимой транскрипции ORF8 SARS-CoV и SARSr-CoV летучих мышей исследовали с использованием репортера люциферазы, 5×ATF6-GL3.В клетках HeLa, транзиентно трансфицированных экспрессионными плазмидами ORF8 вирусов SARSr-CoV летучих мышей Rf1, Rf4092 и WIV1, относительная люциферазная активность репортера 5×ATF6-GL3 повышалась в 5,56–9,26 раз по сравнению с клетками, трансфицированными пустым pCAGGS. вектор, в то время как он был увеличен в 4,42 раза SARS-CoV GZ02 ORF8. В качестве контроля лечение туникамиксином (ТМ) стимулировало транскрипцию примерно в 11 раз (фиг. 9А). Результаты показывают, что различные белки ORF8 вирусов SARSr-CoV летучих мышей могут активировать ATF6, а белки некоторых штаммов обладают более сильным эффектом, чем ORF8 SARS-CoV.
Рис. 9. Функциональная характеристика различных белков ORF8 и ORF8a SARSr-CoV летучих мышей.
(A) Белки ORF8 SARS-CoV и SARSr-CoV летучих мышей индуцируют ATF6-зависимую транскрипционную активность. Клетки HeLa временно трансфицировали экспрессионными плазмидами pcAGGS ORF8 SARS-CoV GZ02, SARSr-CoV Rf1, WIV1 и Rf4092 летучих мышей и репортерной плазмидой 5×ATF6-GL3 в течение 40 часов. Контрольные клетки котрансфицировали репортерной плазмидой и пустым вектором pCAGGS в течение 24 часов и обрабатывали ТМ (2 мкг/мл) или без него в течение дополнительных 16 часов.Клеточные лизаты собирали для двойного анализа люциферазы, и данные представлены как средние значения для трех лунок. (B) Белки ORF8a SARS-CoV и SARSr-CoV летучих мышей запускают апоптоз. Клетки 293T трансфицировали экспрессионными плазмидами ORF8a SARS-CoV Tor2 и SARSr-CoV Rs4084 летучих мышей и векторным контролем pcAGGS в течение 24 часов. Апоптоз анализировали с помощью проточной цитометрии после окрашивания аннексином V и рассчитывали процент апоптотических клеток. Данные показаны как средние значения из трех ячеек.Столбики погрешностей указывают SD. * Р <0,05.
https://doi.org/10.1371/journal.ppat.1006698.g009
Индукция апоптоза с помощью ORF8a недавно идентифицированного SARSr-CoV летучих мышей
Мы провели временную трансфекцию, чтобы проверить, вызывает ли ORF8a SARSr-CoV Rs4084 апоптоз. Как показано на фиг. 9B, апоптозу подверглись 11,76% и 9,40% клеток 293T, трансфицированных экспрессионной плазмидой SARSr-CoV Rs4084-ORF8a и Tor2-ORF8a SARS-CoV, соответственно.Напротив, трансфекция пустым вектором приводила к апоптозу только в 2,79% клеток. Результаты показывают, что Rs4084 ORF8a обладает активностью индукции апоптоза, аналогичной активности SARS-CoV [28].
Обсуждение
Генетически разнообразные SARSr-CoV были обнаружены у различных видов подковоносов в широком географическом ареале в Китае в последнее десятилетие [9–12,14,29]. Однако большинство SARSr-CoV летучих мышей демонстрируют значительную генетическую дистанцию с SARS-CoV, особенно в высоковариабельных областях S1, ORF8 и ORF3 [10,25].Недавно было описано несколько новых SARSr-CoV, которые более тесно связаны с SARS-CoV либо в гене S, либо в ORF8. S-белки RsSHC014, Rs3367, WIV1 и WIV16, о которых сообщалось в наших предыдущих исследованиях, имели от 90% до 97% идентичности аминокислотных последовательностей с белками SARS-CoV человека/циветты [17,18]. Другой штамм из Rhinolophus affinis в Юньнани, названный LYRa11, показал 90% идентичность аминокислотной последовательности с SARS-CoV в гене S [13]. Кроме того, в двух исследованиях было описано 4 новых SARSr-CoV (YNLF_31C/34C и GX2013/YN2013), которые обладали полноразмерной ORF8 со значительно более высоким сходством с таковой у SARS-CoV [22,30].Эти результаты дают убедительные генетические доказательства происхождения SARS-CoV от летучих мышей в отношении гена S или ORF8. Однако все эти SARSr-CoV отличались от SARS-CoV по крайней мере еще одним геном, что позволяет предположить, что ни один из них не был непосредственным предшественником SARS-CoV. Более того, эти SARSr-CoV были обнаружены в популяциях летучих мышей из физически разных мест. Место происхождения истинного прародителя SARS-CoV и эволюционное происхождение SARS-CoV до сих пор оставались неясными. В текущем исследовании мы определили среду обитания летучих мышей, потенциально важную для эволюции SARSr-CoV, где, вероятно, произошла серия событий рекомбинации между различными штаммами SARSr-CoV, что дает новое представление о происхождении SARS-CoV.
SARS впервые появился в провинции Гуандун в конце 2002 г. [7]. Однако SARSr-CoV, обнаруженные на сегодняшний день у летучих мышей из соседних районов провинции Гуандун, показали филогенетические отличия от SARS-CoV, особенно в гене S [9, 10, 14], что позволяет предположить, что SARS-CoV мог произойти из другого региона. Наш анализ филогении SARS-CoV и всех известных SARSr-CoV летучих мышей с использованием последовательности нуклеотидов их неструктурных генов ORF1a и ORF1b, которые составляют большую часть генома, показывает, что эволюция SARSr-CoV сильно коррелирует с их географическим положением. происхождения, а не вида-хозяина.Примечательно, что SARSr-CoV, обнаруженные в провинции Юньнань, более тесно связаны с SARS-CoV, чем штаммы из других регионов Китая. Этот вывод означает, что Юньнань или юго-запад Китая с большей вероятностью является географическим источником SARS-CoV, чем другие регионы Китая, но для подтверждения этого вывода все еще необходимы данные более обширного эпиднадзора.
В нашем лонгитюдном наблюдении за SARSr-CoV в одной пещере в провинции Юньнань, где мы обнаружили Rs3367, RsSHC014, WIV1 и WIV16, распространенность CoV в образцах фекалий варьировалась в зависимости от времени взятия образцов.В целом более высокая распространенность наблюдалась осенью (сентябрь и октябрь), чем весной и в начале лета (апрель и май). Это может быть связано с формированием восприимчивой субпопуляции новорожденных летучих мышей, у которых не выработался собственный иммунитет после родового периода [31]. Еще одним фактором может быть изменение видового состава летучих мышей в пещере в разные сроки отбора проб. Например, в сентябре 2012 года, когда распространенность CoV достигла 51,3%, большинство образцов были из R . sinicus , но в мае 2015 года, когда только 3 из 145 образцов дали положительный результат, Aselliscus stoliczkanus был преобладающим видом летучих мышей в пещере. Нам не удалось амплифицировать последовательности RBD из 15 из 64 положительных образцов SARSr-CoV. Большинство этих образцов имели сравнительно низкую концентрацию вируса (< 10 7 копий/г) (S8 Fig), как показали наши предыдущие количественные исследования [32]. Неудачная амплификация RBD в некоторых образцах с высокой концентрацией вируса, вероятно, была связана с более расходящимися последовательностями в этой области этих геномов SARSr-CoV.
В этой пещере мы получили полноразмерные геномные последовательности еще 11 новых SARSr-CoV от летучих мышей. Наши результаты предполагают совместную циркуляцию различных SARSr-CoV летучих мышей, очень похожих на SARS-CoV в наиболее вариабельных областях S1 (NTD и RBD), ORF8 и ORF3, соответственно, в этом единственном месте. В генах ORF1a, ORF1b, E, M и N вирусы SARSr-CoV, циркулирующие в этой пещере, также имели > 98% идентичности последовательностей аминокислот с SARS-CoV человека/циветты. Таким образом, все строительные блоки генома SARS-CoV присутствовали в SARSr-CoV из этого единственного места в Юньнани в течение периода отбора проб.Кроме того, там также были обнаружены штаммы, тесно связанные с различными репрезентативными SARSr-CoV летучих мышей из других провинций (например, Rs672, HKU3 и Rf1) в регионе RBD. Следовательно, эту пещеру можно рассматривать как богатый генофонд SARSr-CoV летучих мышей, где одновременная циркуляция большого разнообразия штаммов SARSr-CoV привела к необычно разнообразному набору SARSr-CoV.
Во время нашего 5-летнего наблюдения в этой единственной пещере мы впервые сообщили о Rs3367 и WIV1 в 2013 году, причем последовательность RBD очень похожа на последовательность SARS-CoV [17].Совсем недавно мы обнаружили WIV16, который имел RBD, почти идентичный WIV1, но имел гораздо большее сходство с SARS-CoV, чем WIV1, в области NTD S1, что делает его наиболее близким SARSr-CoV к эпидемическим штаммам, идентифицированным на сегодняшний день [18]. В этом исследовании мы обнаружили новый штамм Rs4231 из того же места, который имеет почти идентичную последовательность NTD с WIV16, но отличается от него в RBD, что свидетельствует о рекомбинации. Наш анализ рекомбинации показал, что событие рекомбинации могло иметь место на стыке между кодирующей областью NTD и RBD в геномах Rs4231 и WIV1 и привело к WIV16.Рекомбинация в этом геномном положении также произошла среди других SARSr-CoV, относительно удаленных от SARS-CoV, обнаруженных в этом месте (например, Rs4081 и Rs4247, рис. S5). Частая рекомбинация в этой горячей точке гена S увеличила генетическое разнообразие SARSr-CoV, обитающих в этих популяциях летучих мышей, и, возможно, была ответственна за образование гена S прямого штамма-предшественника SARS-CoV.
Геномы SARS-CoV от пациентов на ранней эпидемической фазе и SARS-CoV циветт содержали одну полноразмерную ORF8 [3,7].Мы обнаружили, что ряд SARSr-CoV летучих мышей из этой пещеры обладал полной ORF8, очень похожей на таковую у раннего SARS-CoV человека/циветты (> 97% идентичности последовательности nt), представленной штаммом Rf4092 (S3C Fig). Это предоставило дополнительные доказательства источника ORF8 SARS-CoV человека у летучих мышей [22,30]. Напротив, ORF8 была разделена на перекрывающиеся ORF8a и ORF8b в большинстве человеческих штаммов SARS-CoV от пациентов с более поздней фазой из-за приобретения делеции 29 нуклеотидов [8,26]. В этом исследовании мы впервые обнаружили SARSr-CoV летучих мышей с ORF8a и ORF8b, очень похожими на более поздние фазы SARS-CoV человека, хотя разделение ORF8 в SARSr-CoV летучих мышей и SARS-CoV человека были двумя независимыми событиями.Наш анализ рекомбинации предполагает, что этот штамм, Rs4084, вероятно, получил свою ORF8 от Rf4092 посредством рекомбинации, за которой последовало развитие 5-нуклеотидной делеции, которая привела к расщеплению. Это предполагает, что область ORF8 в геномах SARSr-CoV летучих мышей склонна к делециям, как и в случае SARS-CoV человека [3,25]. Наконец, анализ рекомбинации предполагает, что наследственный штамм SARS-CoV SZ3 мог бы образоваться, если бы рекомбинация вокруг ORF8 произошла между линиями, которые привели к WIV16 и Rf4092.В совокупности данные о событиях рекомбинации среди SARSr-CoV, населяемых летучими мышами в этом единственном месте, позволяют предположить, что прямой предшественник SARS-CoV мог возникнуть в результате серии рекомбинаций внутри гена S и вокруг ORF8. За этим мог последовать перенос от летучих мышей к циветтам и людям либо в регионе, либо во время перемещения инфицированных животных через торговлю дикими животными. Однако, учитывая скудость данных о торговле животными до вспышки атипичной пневмонии, вероятную высокую географическую погрешность выборки при наблюдении за летучими мышами на наличие SARSr-CoV на юге Китая, а также возможность того, что в других пещерах обитают сходные сообщества видов летучих мышей и богатое разнообразие атипичной пневмонии. -CoVs, на данный момент нельзя сделать определенный вывод о географическом происхождении SARS-CoV.
Р . sinicus считаются первичными естественными хозяевами SARS-CoV, поскольку все SARSr-CoV, высоко гомологичные SARS-CoV по S-гену, были преимущественно обнаружены у этого вида. Однако отмечается, что ранее сообщалось о двух SARSr-CoV от R . ferrumequinum показал самое близкое филогенетическое положение к SARS-CoV в деревьях ORF1a/1b. Эти штаммы были обнаружены в другом месте в провинции Юньнань, в 80 км от пещеры, обследованной в настоящем исследовании [22].Эта информация также подтверждает предположение о том, что SARS-CoV мог возникнуть в этом регионе. Тем не менее, поскольку корреляция между видами-хозяевами и филогении ORF1ab SARSr-CoV кажется ограниченной, необходимо получить больше последовательностей SARSr-CoV от различных видов летучих мышей Rhinolophus как в Юньнани, так и в других местах на юге Китая. В частности, будет важно оценить, соответствует ли R . ferrumequinum сыграл более важную роль в эволюции ORF1ab SARS-CoV.
Изученная нами пещера находится примерно в 60 км от города Куньмин. Помимо ряда видов ринолофидов и гиппозидеридов, у которых были обнаружены SARSr-CoV, там также присутствовали другие летучие мыши, такие как миотисы. Температура в пещере около 22–25°C, а влажность около 85–90%. Физическая природа пещеры не уникальна, но, похоже, в репродуктивный период в ней обитает особенно плотная популяция летучих мышей. Подобные пещеры, в которых живут популяции летучих мышей разных видов, не редкость и в других районах Юньнани.Мы предполагаем, что усилия по изучению экологии, разнообразия видов-хозяев и популяций вирусных штаммов в этих пещерах могут предоставить важную информацию о том, что движет эволюцией SARSr-CoV.
Наши предыдущие исследования продемонстрировали способность как WIV1, так и WIV16 использовать ортологи ACE2 для проникновения в клетки и эффективно реплицироваться в клетках человека [17,18]. В этом исследовании мы подтвердили использование человеческого ACE2 в качестве рецептора двух новых SARSr-CoV, используя химерные вирусы с заменой остова WIV1 на ген S недавно идентифицированных SARSr-CoV.Белок S Rs7327 отличался от белка WIV1 и WIV16 тремя аминокислотными остатками в мотиве связывания с рецептором, включая один контактный остаток (484 аминокислотных остатка) с человеческим ACE2. Это различие, по-видимому, не влияло на его проникновение и эффективность репликации в клетках, экспрессирующих ACE2 человека. Предыдущее исследование с использованием инфекционного клона SARS-CoV показало, что S-белок RsSHC014 может эффективно использовать человеческий ACE2 [33], несмотря на то, что он отличается от SARS-CoV и WIV1 в RBD (рис. S1). Мы исследовали инфекционность Rs4231, который имеет схожую последовательность RBD с RsSHC014, но имеет отличную последовательность NTD, и обнаружили, что химерный вирус WIV1-Rs4231S также легко реплицируется в клетках HeLa, экспрессирующих молекулу ACE2 человека.Новый живой SARSr-CoV, который мы выделили в текущем исследовании (Rs4874), имеет ген S, почти идентичный гену WIV16. Как и ожидалось, он также способен использовать человеческий ACE2. Эти результаты показывают, что различные варианты S-белка SARSr-CoV без делеций в их RBD способны использовать человеческий ACE2. Напротив, наше предыдущее исследование показало, что белок S R . sinicus SARSr-CoV с делециями (Rp3) не смог использовать ACE2 человека, циветты и летучей мыши для проникновения в клетку [34]. В этом исследовании, в дополнение к Rs4231 и Rs7327, мы также сконструировали инфекционные клоны с геном S Rs4081, Rf4075, Rs4085, Rs4235 и As6526, которые все содержали делеции в их RBD.Эти 7 штаммов, а также Rs4874 и ранее изученные WIV1 и RsSHC014 могут представлять все типы S-вариантов SARSr-CoV в этом месте (рис. S3A). Однако ни один из штаммов с делециями в RBD не удалось выделить из клеток Vero E6. Таким образом, две разные клады гена S SARSr-CoV могут представлять использование разных рецепторов у летучих мышей-хозяев.
Полноразмерный белок ORF8 SARS-CoV представляет собой белок, ассоциированный с мембраной просветного эндоплазматического ретикулума (ER), который индуцирует активацию ATF6, фактора транскрипции, регулируемого стрессом ER, который активирует транскрипцию шаперонов ER, участвующих в фолдинге белка [35 ].Мы амплифицировали гены ORF8 Rf1, Rf4092 и WIV1, которые представляют три разных генотипа ORF8 SARSr-CoV летучих мышей (S3C Fig), и сконструировали экспрессионные плазмиды. Все три белка ORF8, транзиентно экспрессируемые в клетках HeLa, могут стимулировать ATF6-зависимую транскрипцию. Среди них ORF8 WIV1, которая сильно отличается от ORF8 SARS-CoV, продемонстрировала самую сильную активацию. Результаты показывают, что варианты белков ORF8 SARSr-CoV летучих мышей могут играть роль в модуляции стресса ER путем активации пути ATF6.Кроме того, было показано, что белок ORF8a SARS-CoV поздней фазы индуцирует апоптоз [28]. В этом исследовании мы обнаружили, что белок ORF8a недавно идентифицированного SARSr-CoV Rs4084, который содержал вставку из 8 аминокислот по сравнению с ORF8a SARS-CoV, также значительно запускал апоптоз в клетках 293T.
По сравнению с ORF3b из 154 аминокислотных остатков SARS-CoV белки ORF3b всех ранее идентифицированных SARSr-CoV летучих мышей были меньше по размеру из-за ранней терминации трансляции.Однако мы впервые обнаружили ORF3b без С-концевого укорочения у SARSr-CoV летучей мыши, Rs7327, которая отличалась от ORF 3b штамма SARS-CoV GZ02 только одним аминокислотным остатком. ORF3b SARS-CoV противодействует функции интерферона, модулируя активность регуляторного фактора 3 IFN (IRF3) [27]. Как предполагалось в предыдущих исследованиях, С-конец, содержащий сигнал ядерной локализации, может не требоваться для активности антагониста IFN ORF3b [36]. Наши предыдущие исследования также показали, что белок ORF3b SARSr-CoV летучей мыши, названный Rm1, который был укорочен на С-конце до 56 а.о. и имел 62% идентичность последовательности а.о. с SARS-CoV, по-прежнему проявлял активность антагониста IFN [37].В дальнейших исследованиях очень интересно выяснить, проявляют ли ORF3b Rs7327 и другие версии укороченной ORF3b, такие как WIV1 и WIV16, также профили антагонизма к IFN.
В целом наши результаты 5-летнего продольного исследования убедительно демонстрируют, что все строительные блоки генома пандемического SARS-CoV присутствуют в SARSr-CoV летучих мышей из одного места в провинции Юньнань. Данные показывают, что среди этих SARSr-CoV в одной и той же пещере происходили частые события рекомбинации. Хотя мы не можем исключить возможность того, что аналогичные генофонды SARSr-CoV существуют где-то еще, мы предоставили достаточно доказательств, чтобы сделать вывод, что SARS-CoV, скорее всего, произошел от подковоносых летучих мышей в результате рекомбинации среди существующих SARSr-CoV.Кроме того, мы также обнаружили, что различные SARSr-CoV, способные использовать человеческий ACE2, все еще циркулируют среди летучих мышей в этом регионе. Таким образом, возможен риск распространения на людей и возникновения заболевания, похожего на ОРВИ. Это особенно важно, учитывая, что ближайшая деревня к обследованной нами пещере летучих мышей находится всего в 1,1 км, что указывает на потенциальный риск контакта с летучими мышами для местных жителей. Таким образом, мы предлагаем продолжить мониторинг эволюции SARSr-CoV в этом и других местах, а также изучение поведенческого риска инфицирования человека и серологические обследования людей, чтобы определить, происходит ли распространение в этих местах, и разработать стратегии вмешательства. во избежание появления болезней в будущем.
Материалы и методы
Заявление об этике
Все процедуры отбора проб проводились ветеринарами с одобрения Комитета по этике животных Уханьского института вирусологии (WIVH05210201). Исследование проводилось в соответствии с Руководством по уходу и использованию диких млекопитающих в исследованиях Китайской Народной Республики.
Отбор проб
Отбор проб летучих мышей был проведен десять раз с апреля 2011 г. по октябрь 2015 г. в разные сезоны в их естественной среде обитания в одном месте (пещере) в Куньмине, провинция Юньнань, Китай.Все члены полевых бригад носили соответствующие средства индивидуальной защиты, в том числе маски N95, непроницаемые перчатки, одноразовую верхнюю одежду и защитные очки. Летучих мышей отлавливали, а образцы фекалий собирали, как описано ранее [9]. Чистые пластиковые листы размером 2,0 на 2,0 м помещали под известные места ночевок летучих мышей примерно в 18:00 каждый вечер для сбора образцов фекалий. Свежие фекальные шарики собирали с простыней рано утром следующего дня. Каждый образец (приблизительно 1 грамм фекального шарика) собирали в 1 мл вирусной транспортной среды, состоящей из сбалансированного солевого раствора Хенкса при pH 7.4, содержащие БСА (1%), амфотерицин (15 мкг/мл), пенициллин G (100 ЕД/мл) и стрептомицин (50 мкг/мл), и хранили при -80°С до обработки. Летучие мыши, пойманные в ловушку для этого исследования, были выпущены обратно в их среду обитания.
Экстракция РНК, ПЦР-скрининг и секвенирование
Образцы фекальных мазков или гранул встряхивали в течение 1 минуты, и из каждого образца отбирали 140 мкл супернатанта после центрифугирования при 3000 об/мин при 4°C в течение 1 минуты. Вирусную РНК экстрагировали с помощью набора Viral RNA Mini Kit (Qiagen) в соответствии с инструкциями производителя.РНК элюировали 60 мкл буфера AVE (вода без РНКаз с 0,04% азида натрия, Qiagen), разделяли на аликвоты и хранили при -80°С. Одноэтапная полугнездовая ОТ-ПЦР (Invitrogen) использовалась для обнаружения присутствия последовательностей коронавируса, как описано ранее, с использованием набора праймеров, нацеленных на 440-нуклеотидный фрагмент в гене РНК-зависимой РНК-полимеразы ( RdRp ) все известные альфа- и бета-коронавирусы [20]. Для первого раунда ПЦР 25 мкл реакционной смеси содержали 12,5 мкл ПЦР 2 × буфер для реакционной смеси, 10 пмоль каждого праймера, 2.5 мМ MgSO 4 , 20 ед. ингибитора РНКазы, 1 мкл смеси ферментов SuperScript III/Platinum Taq и 5 мкл матрицы РНК. Амплификацию проводили следующим образом: 50°С в течение 30 мин, 94°С в течение 2 мин, затем 40 циклов, состоящих из 94°С в течение 15 с, 52°С в течение 30 с, 68°С в течение 40 с и окончательное удлинение 68°С в течение 5 мин. Для второго раунда ПЦР 25 мкл реакционной смеси содержали 2,5 мкл реакционного буфера для ПЦР, 5 пмоль каждого праймера, 50 мМ MgCl 2 , 0,5 мМ dNTP, 0,1 мкл Platinum Taq Enzyme (Invitrogen) и 1 мкл продукта первого круглый ПКР.Амплификацию проводили следующим образом: 94°С в течение 3 мин, затем 35 циклов, состоящих из 94°С в течение 30 с, 52°С в течение 30 с, 72°С в течение 40 с и заключительного удлинения 72°С в течение 7 мин. Участок RBD амплифицировали с использованием описанного ранее одноэтапного метода вложенной ОТ-ПЦР [17].
продукта ПЦР очищали в геле и секвенировали на ДНК-анализаторе ABI Prism 3730 (Applied Biosystems, США). Продукты ПЦР с низкой концентрацией или вызывающие гетерогенность на хроматограммах секвенирования клонировали в pGEM-T Easy Vector (Promega) для секвенирования.Положительные образцы в этом исследовании были названы с использованием сокращенного названия вида летучих мышей плюс идентификационный номер образца (например, Rs4081). Чтобы подтвердить вид летучей мыши в отдельном образце, была проведена ПЦР-амплификация гена цитохрома b ( Cytob ) или субъединицы 1 НАДН-дегидрогеназы ( ND1 ) с использованием ДНК, выделенной из фекалий или мазков [38,39].
Секвенирование полноразмерных геномов
Полные геномные последовательности 11 SARSr-CoV были определены с помощью одноэтапной ПЦР (Invitrogen) амплификации перекрывающихся геномных фрагментов с вырожденными праймерами, разработанными путем множественного выравнивания доступных последовательностей SARS-CoV и SARSr-CoV летучих мышей, депонированных в GenBank, и дополнительных специфических праймеров разработан на основе результатов предыдущих раундов секвенирования в этом исследовании.Последовательности праймеров доступны по запросу. Последовательности 5’- и 3’-концов генома были получены с помощью 5’- и 3’-RACE (Roche) соответственно. Продукты ПЦР ожидаемого размера очищали в геле и подвергали непосредственному секвенированию. Каждый фрагмент секвенировали не менее двух раз. Хроматограмма секвенирования каждого продукта была тщательно изучена, и гетерогенность последовательности не наблюдалась. Для некоторых фрагментов с низкой концентрацией ампликонов продукты ПЦР клонировали в pGEM-T Easy Vector (Promega) для секвенирования.По меньшей мере пять независимых клонов секвенировали для получения консенсусной последовательности. Совместное присутствие последовательностей различных SARSr-CoV не было обнаружено ни в одном из ампликонов. Последовательности перекрывающихся геномных фрагментов были собраны для получения полноразмерных последовательностей генома, причем каждая перекрывающаяся последовательность длиннее 100 п.н.
Анализ эволюции
Полноразмерные геномные последовательности 15 SARSr-CoV, обнаруженных у летучих мышей в пещере, обследованной в этом исследовании, были сопоставлены с последовательностями отдельных SARS-CoV с помощью MUSCLE [40].Выровненные последовательности сканировали на наличие событий рекомбинации с помощью программы обнаружения рекомбинации (RDP) [41]. Потенциальные события рекомбинации, предполагаемые сильными значениями P (<10 -20 ), были дополнительно подтверждены с использованием графика сходства и анализа начальной загрузки, реализованного в Simplot 3.5.1 [42]. Филогенетические деревья на основе нуклеотидных последовательностей были построены с использованием алгоритма максимального правдоподобия в модели LG со значениями начальной загрузки, определенными по 1000 повторов в PhyML (версия 3.0) пакет программ [43].
Выделение вируса
Клеточная линия Vero E6 была любезно предоставлена Австралийской лабораторией здоровья животных, CSIRO (Джилонг, Австралия). Монослой Vero E6 поддерживали в среде DMEM с добавлением 10% эмбриональной телячьей сыворотки (FCS). Образцы фекалий (в 200 мкл буфера) центрифугировали в градиенте при 3000–12000 g, а супернатант разбавляли 1:10 в DMEM перед добавлением к клеткам Vero E6. После инкубации при 37°С в течение 1 ч инокулят удаляли и заменяли свежей средой DMEM с 2% FCS.Клетки инкубировали при 37°С и ежедневно проверяли на цитопатический эффект. Во все среды для культивирования тканей добавляли тройные антибиотики пенициллин/стрептомицин/амфотерицин (Gibco) (пенициллин 200 МЕ/мл, стрептомицин 0,2 мг/мл, амфотерицин 0,5 мкг/мл). Для каждого образца проводили три слепых пассажа. После каждого пассажа культуральный супернатант и клеточный осадок исследовали на наличие SARSr-CoV с помощью ОТ-ПЦР с использованием специфических праймеров, нацеленных на ген RdRp или S. Вирусы, которые вызывали выраженный цитопатический эффект и могли быть обнаружены в трех слепых пассажах с помощью ОТ-ПЦР, были дополнительно подтверждены с помощью электронной микроскопии.
Конструирование рекомбинантных вирусов
Рекомбинантные вирусы с геном S нового SARSr-CoV летучих мышей и основой инфекционного клона SARSr-CoV WIV1 были сконструированы с использованием обратной генетической системы, описанной ранее [23] (S9 Fig). Фрагменты E и F реамплифицировали с парами праймеров (FE, 5′-AGGGCCCACCTGGCACTGGTAAGAGTCATTTTGC-3′, R-EsBsaI, 5′-ACTGGTCTCTTCGTTTAGTTATTAACTAAAATATCACTAGACACC-3′) и (F-FsBsaI, 5′-TGAGGTCTCCGAACTTATGGATTTGTTTATGAG-3′, RF , 5′-AGGTAGGCCTCTAGGGCAGCTAAC-3′) соответственно.Продукты были названы фрагментами Es и Fs, которые оставляют область, кодирующую ген спайка, как независимый фрагмент. Сайты BsaI (5’-GGTCTCN|NNNN-3’) вводили в 3’-конец фрагмента Es и 5’-конец фрагмента Fs соответственно. Шипообразную последовательность Rs4231 амплифицировали с парой праймеров (F-Rs4231-BsmBI, 5’-AGTCGTCTCAACGAACATGTTTTATTTTCTTATTCTTTCTCACTCTCAC-3’ и R-Rs4231-BsmBI, 5’-TCACGTCTCAGTTCGTTTATGTGTAATGTAATTTGACACCCTTG-3’). Последовательность гена S Rs7327 амплифицировали с парой праймеров (F-Rs7327-BsaI, 5’-AGTGGTCTCAAACCAACTGAAATTGTTAGTTTTAGTTTTTGCTAC-3’ и R-Rs7327-BsaI, 5’-TCAGGTCTCAGTTCGTTTATGTGTAATGTAATTTAACACCCTTG-3’).Фрагменты Es и Fs расщепляли BglI (NEB) и BsaI (NEB). Ген Rs4231 S расщепляли с помощью BsmBI. Ген Rs7327 S расщепляли с помощью BsaI. Другие фрагменты и бактериальную искусственную хромосому (ВАС) готовили, как описано ранее. Затем два подготовленных фрагмента спайковой ДНК по отдельности встраивали в ВАС вместе с Es, Fs и другими фрагментами. Были проверены правильные инфекционные клоны BAC. Химерные вирусы были спасены, как описано ранее [23].
Определение инфекционности вируса с помощью иммунофлуоресцентного анализа
Клеточная линия HeLa была любезно предоставлена Австралийской лабораторией здоровья животных, CSIRO (Джилонг, Австралия).Клетки HeLa, экспрессирующие человеческий ACE2, конструировали, как описано ранее [17]. Клетки HeLa, экспрессирующие клетки ACE2 человека и Vero E6, культивировали на покровных стеклах в 24-луночных планшетах (Corning) и инкубировали с недавно выделенными или рекомбинантными SARSr-CoV летучих мышей при множественности заражения (MOI) = 1,0 в течение 1 часа. Инокулят удаляли, клетки дважды промывали PBS и добавляли среду. В качестве отрицательного контроля использовали клетки Vero E6 без инокуляции вируса и клетки HeLa без ACE2. Через 24 часа после инфицирования клетки промывали PBS и фиксировали 4% формальдегидом в PBS (рН 7.4) при 4°С в течение 20 мин. Экспрессию ACE2 определяли с использованием козьего иммуноглобулина против человеческого ACE2, а затем меченного FITC иммуноглобулина осла против козьего иммуноглобулина (PTGLab). Репликация вируса была обнаружена с использованием кроличьего антитела против нуклеокапсидного белка SARSr-CoV летучей мыши Rp3, а затем Cy3-конъюгированного мышиного антикроличьего IgG. Ядра окрашивали DAPI. Окрашивание наблюдали под конфокальным микроскопом FV1200 (Olympus).
Определение репликации вируса в клетках Vero E6 методом анализа бляшек
клетки Vero E6 были инфицированы вирусами WIV1, Rs4874, WIV1-Rs4231S и WIV1-Rs7327S при множественности заражения 1.0 и 0,01. После инкубации в течение часа клетки трижды промывали DHanks и снабжали DMEM, содержащей 2% FCS. Образцы собирали через 0, 10, 27 и 48 часов после инфицирования. Титры вируса определяли анализом бляшек.
Определение репликации вируса в клетках HeLa, экспрессирующих человеческий ACE2, с помощью количественной ОТ-ПЦР
клеток HeLa, экспрессирующих человеческий ACE2, инокулировали WIV1, Rs4874, WIV1-Rs4231S и WIV1-Rs7327S при множественности заражения 1,0 и инкубировали в течение 1 часа при 37°C.После удаления инокулята к клеткам добавляли среду, содержащую 1% FBS. Супернатанты собирали через 0, 12, 24 и 48 часов. Титры вируса определяли с помощью количественной ОТ-ПЦР, нацеленной на частичный ген N, со стандартной кривой, которая выражает корреляцию между значением Ct и титром вируса (показанным как TCID50/мл). Стандартную кривую строили с использованием разведений РНК из очищенного штамма вируса Rs4874 (с титром 2,15 × 10 6 TCID50/мл). Для количественной ПЦР РНК экстрагировали из 140 мкл каждого супернатанта с помощью набора Viral RNA Mini Kit (Qiagen) в соответствии с инструкциями производителя и элюировали в 60 мкл буфера AVE.ПЦР проводили с помощью набора TaqMan AgPath-ID One-Step RT-PCR Kit (Applied Biosystems) в 25 мкл реакционной смеси, содержащей 4 мкл РНК, 1 × смесь ферментов ОТ-ПЦР, 1 × буфер ОТ-ПЦР, 40 пмоль вперед. праймер (5′-GTGGTGGTGACGGCA AAATG-3′), обратный праймер 40 пмоль (5′-AAGTGAAGCTTCTGGGCCAG-3′) и зонд 12 пмоль (5′-FAM-AAAGAGCTCAGCCCCAGATG-BHQ1-3′). Амплификацию проводили следующим образом: 50°С в течение 10 мин, 95°С в течение 10 мин, затем 50 циклов, состоящих из 95°С в течение 15 с и 60°С в течение 20 с.
Плазмиды
Гены ORF8 SARSr-CoV WIV1 и Rf4092 летучих мышей и ген ORF8a SARSr-CoV летучих мышей Rs4084 были амплифицированы с помощью ПЦР из вирусной РНК, выделенной из выделенного вируса или образцов фекалий.Ген ORF8 SARS-CoV GZ02 и SARSr-CoV Rf1 летучих мышей, а также ген ORF8a SARS-CoV Tor2 были синтезированы компанией Tsingke Biological Technology Co., Ltd (Ухань, Китай). Все гены были клонированы в вектор pCAGGS, сконструированный с С-концевой НА-меткой. Экспрессию белков подтверждали вестерн-блоттингом с использованием mAb против метки HA. Пять тандемных копий консенсусных сайтов связывания ATF6 были синтезированы и вставлены в вектор pGL3-Basic для конструирования репортерной плазмиды люциферазы 5×ATF6-GL3, в которой ген люциферазы находится под контролем минимального промотора c- fos и консенсусные сайты связывания ATF6.
Репортерный анализ люциферазы
клеток HeLa в 24-луночных планшетах трансфицировали с использованием реагента Lipofectamine 3000 (Life Technologies) в соответствии с инструкциями производителя. Клетки на лунку котрансфицировали 600 нг репортерной плазмиды 5×ATF6-GL3, 300 нг каждой экспрессионной плазмиды SARS-CoV и SARSr-CoV ORF8 или пустого вектора и 20 нг pRL-TK (Promega), которые служили в качестве внутренний контроль. Клетки инкубировали в течение 24 часов и обрабатывали 2 мкг/мл туникамицина или без него в течение 16 часов.Клетки собирали и лизировали. Активность люциферазы определяли с использованием системы анализа двойной люциферазы (Promega). Эксперимент проводили в трехкратной повторности лунок.
Количественное определение апоптотических клеток
Клетки293T в 12-луночных планшетах трансфицировали с использованием реагента Lipofectamine 3000 (Life Technologies) в соответствии с инструкциями производителя. Клетки на лунку трансфицировали 3 мкг экспрессионной плазмиды SARS-CoV Tor2 или SARSr-CoV Rs4084 ORF8a или пустого вектора.Через 24 часа после трансфекции апоптотические клетки определяли количественно с использованием набора для обнаружения апоптоза аннексина V-флуоресцеина (FITC)/PI (Yeasen Biotech, Шанхай) в соответствии с инструкциями производителя. Апоптоз анализировали с помощью проточной цитометрии. Эксперимент проводили в трехкратной повторности лунок.
Регистрационные номера
Полные геномные последовательности штаммов As6526, Rs4081, Rs4084, Rf4092, Rs4231, Rs4237, Rs4247, Rs4255, Rs4874, Rs7327 и Rs9401, связанных с атипичной пневмонией у летучих мышей, были депонированы в базе данных GenBank под номерами доступа от KY417172 до 2KY41 соответственно. .
Проспективное, рандомизированное, многоцентровое исследование внутрикостной абляции базивертебрального нерва для лечения хронической боли в пояснице ), которые не поддаются адекватному лечению при стандартной помощи. Предыдущие 2-летние данные группы лечения ложно-контролируемого рандомизированного контролируемого исследования (РКИ) показали сохранение клинических улучшений через 2 года после радиочастотной (РЧ) аблации базивертебрального нерва (БВН).
ЦЕЛЬ
Целью данного РКИ было сравнить эффективность внутрикостной РЧ-абляции БВН со стандартным лечением хронической БВП в конкретной подгруппе пациентов с подозрением на вертеброгенную симптоматику.
ДИЗАЙН/УСТАНОВКИ ИССЛЕДОВАНИЯ
Проспективное параллельное открытое РКИ было проведено в 20 центрах США.
ОБРАЗЕЦ ПАЦИЕНТА
В общей сложности 140 пациентов с хронической БН продолжительностью не менее 6 месяцев, с изменениями замыкательной пластинки позвонков типа Modic 1 или 2 между L3 и S1, были рандомизированы 1:1 для проведения либо РЧ-абляции БВН, либо продолжения лечения. стандартный уход.
ПОКАЗАТЕЛИ РЕЗУЛЬТАТОВ
Индекс нетрудоспособности Освестри (ODI) собирали на исходном уровне, через 3, 6, 9 и 12 месяцев после процедуры. Вторичные показатели исхода включали 10-балльную визуально-аналоговую шкалу (ВАШ) для LBP, ODI и VAS, показатели респондентов, SF-36 и EQ-5D-5L. Первичной конечной точкой было межгрупповое сравнение среднего изменения ODI от исходного уровня до 3 месяцев после лечения.
МЕТОДЫ
Пациенты были рандомизированы в соотношении 1:1 для получения радиочастотной абляции или продолжения стандартного лечения.Результаты, о которых пациенты сообщали сами, собирались с использованием утвержденных вопросников при каждом визите в рамках исследования. Промежуточный анализ для оценки превосходства был предварительно определен и контролировался независимым комитетом по управлению данными, когда минимум 60% пациентов завершили свое 3-месячное посещение по первичной конечной точке.
РЕЗУЛЬТАТЫ
Промежуточный анализ показал явное статистическое превосходство (p<0,001) для всех первичных и вторичных исходов, сообщаемых пациентами, в группе радиочастотной аблации по сравнению со стандартной группой лечения.Это привело к рекомендации комитета по управлению данными прекратить регистрацию в исследовании и предложить ранний переход в контрольную группу. Эти результаты состоят из исходов 104 пациентов, включенных в анализ намерения лечить 3-месячную первичную конечную точку, которая включала 51 пациента в группе радиочастотной аблации и 53 пациента в группе стандартного лечения. Исходный ODI составил 46,1, ВАШ — 6,67, средний возраст — 50 лет. Процент пациентов с симптомами БНС ≥5 лет составил 67,3%. При сравнении группы РЧ-абляции со стандартной терапией средние изменения ODI через 3 месяца составили -25.3 балла против -4,4 балла соответственно, в результате чего скорректированная разница составила 20,9 балла (p<0,001). Средние изменения по ВАШ составили -3,46 против -1,02 соответственно, скорректированная разница 2,44 см (p<0,001). В группе РЧ-абляции 74,5% пациентов достигли улучшения ODI на ≥10 баллов по сравнению с 32,7% в группе стандартного лечения (p<0,001).
ВЫВОДЫ
Минимально инвазивная радиочастотная абляция БВН привела к значительному уменьшению боли и улучшению функции через 3 месяца у пациентов с хронической вертеброгенной БНС.