
Судебная практика ст 619 гк рф — Народный интерес
Ст. 619 ГК РФ: судебная практика
К отношениям, которые касаются аренды, применяются правила гл. 34 ГК, если иное не предусматривается специальными нормами. В соответствии с общим порядком, субъекты заключают соглашение, по которому одна сторона передает, а вторая принимает имущество на установленный срок в пользование и владение. При этом последняя выплачивает за это определенную сумму. Она устанавливается соглашением сторон. Правила прекращения правоотношений, связанных с возмездным использованием вещи, определены в ст. 619,620 ГК РФ.
Особенности сделки
Арендатор в процессе эксплуатации объекта, переданного ему, может получать доход, плоды или создавать продукцию. Они являются его собственностью. Предметом арендного договора выступает индивидуально-определенная вещь, которая не теряет своих свойств в процессе использования по назначению. Одним из обязательных требований при заключении соглашения выступает точное описание передаваемого имущества.
Субъектный состав
В качестве сторон сделки выступают собственник или иной законный владелец (арендодатель) и пользователь (арендатор). Участниками отношений могут быть физические или юрлица, а также государство, реализующее свои права через уполномоченные органы. Арендодатель – собственник/законный владелец или лицо, управомоченное им/законом. Субъект, у которого в хозяйственном ведении находится движимая вещь, может передавать ее в пользование иным лицам без согласия титульного обладателя. Арендатором, по общему правилу, может являться любой субъект. Однако в ряде случаев законодательство может предусматривать ограничения. Например, в договоре аренды организации или лизинговом соглашении оба участника должны быть предпринимателями (юрлицами). Если вещь предоставляется в прокат, одна сторона должна быть физлицом.
Ст. 619 ГК РФ: расторжение договора аренды
Законодательство предусматривает ряд случаев, в которых отношения между субъектами могут быть прекращены досрочно.
- Эксплуатирует имущество с существенными нарушениями условий соглашения либо неоднократным их допущением.
- В значительной степени ухудшает состояние вещи.
- Допустил неуплату суммы, установленной за эксплуатацию объекта, более двух раз подряд по истечении предусмотренного соглашением срока.
- Не осуществляет капремонт имущества в установленный договором период. Если сроки в соглашении не прописаны, то этот пункт нормы 619 ГК РФ действует в случаях, когда законодательство, иной нормативный акт, условия сделки предусматривают необходимость проведения ремонтных работ в разумные временные промежутки.
Стороны могут предусмотреть и иные основания для прекращения правоотношений досрочно. Арендодатель может требовать расторжения договора до окончания установленного в нем периода только после направления пользователю письменного уведомления о необходимости выполнения последним принятых обязательств.
Ст. 619 ГК РФ с комментариями
Прекращение правоотношений, связанных с использованием имущества за плату, допускается в случаях, установленных законодательством, прочими нормами или соглашением. Статья 619 ГК РФ предусматривает 4 основания. Первым является эксплуатация имущества с допущением нарушений. Они могут быть неоднократными или существенными. Последними считаются такие нарушения, которые влекут для другого участника такой ущерб, что при его возникновении субъект в значительной мере лишается того, на что он мог рассчитывать при совершении сделки. Ст. 619 ГК РФ действует, например, в случае, когда пользователь передал объект в перенаем без согласия собственника.
Ухудшение состояния вещи
По смыслу нормы 619 ГК РФ, под таким действием следует понимать поведенческий акт, вследствие которого объект становится неисправным. Другими словами, его состояние ухудшено до степени, превышающей нормальный износ. Это, как правило, обуславливается невыполнением пользователем обязанности по поддержанию вещи в исправном состоянии, выполнению текущего ремонта, несению расходов на ее содержание. Предъявляя требования в рамках нормы 619 ГК РФ , собственник/законный владелец должен доказать, что степень полезности свойств объекта в ходе его эксплуатации настолько снизилась, что это повлекло ущерб, при котором он лишается того, на что рассчитывал при совершении сделки.
Это, как правило, обуславливается невыполнением пользователем обязанности по поддержанию вещи в исправном состоянии, выполнению текущего ремонта, несению расходов на ее содержание. Предъявляя требования в рамках нормы 619 ГК РФ , собственник/законный владелец должен доказать, что степень полезности свойств объекта в ходе его эксплуатации настолько снизилась, что это повлекло ущерб, при котором он лишается того, на что рассчитывал при совершении сделки.
Уклонение от внесения платы
По смыслу нормы 619 ГК РФ, нарушением условий соглашения будет выступать просрочка погашения денежного обязательства не менее трех раз подряд. В такой ситуации собственник может потребовать досрочного внесения платы. При этом он сам устанавливает срок для этого, если иное не определено в соглашении. Собственник не может требовать внесения платы больше чем за 2 раза подряд. Если пользователь продолжает уклоняться от исполнения обязательства, кредитор вправе расторгнуть договор.
Собственник может требовать досрочного расторжения соглашения и в том случае, если задолженность по оплате была погашена арендатором. Между тем законодательство ограничивает данное право. Если собственник не предъявит требование о расторжении соглашения в разумный срок после погашения арендатором возникшей задолженности, он теряет данную возможность. Данное положение подтверждается Постановлением Пленума ВАС № 73 от 17.11.2011 г. (п. 23, абзац 2).
Между тем законодательство ограничивает данное право. Если собственник не предъявит требование о расторжении соглашения в разумный срок после погашения арендатором возникшей задолженности, он теряет данную возможность. Данное положение подтверждается Постановлением Пленума ВАС № 73 от 17.11.2011 г. (п. 23, абзац 2).
Обязательства по проведению капремонта
Они могут предусматриваться непосредственно в арендном соглашении или в законодательстве. Кроме этого, стороны вправе установить срок, в который пользователь должен провести капремонт. Если он не определен, то в расчет принимается разумный период. Им считается срок, который необходим для поддержания вещи в нормальном, пригодном к использованию состоянии.
В соответствии с п. 25 Обзора Президиума ВАС № 66, в перечень оснований могут включаться и иные случаи. При этом список не может быть сокращен по договоренности сторон. Законодательство допускает включение в соглашение оснований, не связанных с нарушениями обязательств.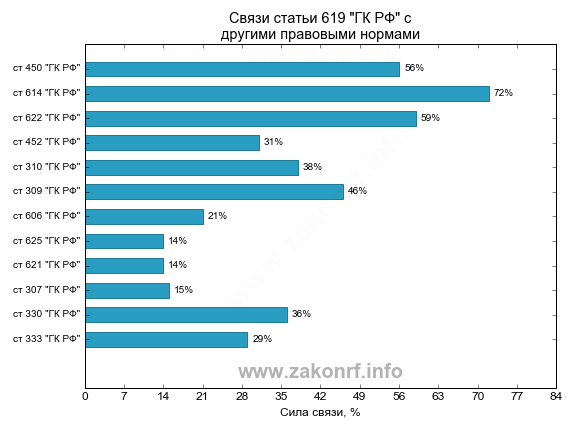 Например, в судебной практике имеют место случаи расторжения договора по требованию собственника в связи с отнесением эксплуатируемого имущества в перечень реконструируемых объектов, производственной необходимостью использовать его самим законным владельцем и так далее.
Например, в судебной практике имеют место случаи расторжения договора по требованию собственника в связи с отнесением эксплуатируемого имущества в перечень реконструируемых объектов, производственной необходимостью использовать его самим законным владельцем и так далее.
Расторжение договора может осуществляться в судебном порядке по основаниям, не указанным в ст. 619, если инстанция признает их существенными. Так, Президиум ВАС указывает, что в качестве таких обстоятельств могут выступать невнесение платы однократно или неполная выплата установленной суммы. В данном случае суд оценивает существенность нарушений.
Расторжение договора включает в себя 2 этапа. На первом этапе заинтересованное лицо обязано соблюсти досудебный порядок. Такая необходимость прямо устанавливается в норме 619 ГК РФ. Досудебный порядок предполагает направление уведомления пользователю с сообщением о необходимости исполнить принятое обязательство в разумные сроки. Если арендатор не предпринял соответствующих мер для удовлетворения претензий, арендодатель направляет письменное уведомление о расторжении договора. Срок для получения ответа по общему правилу составляет 30 дней.
Срок для получения ответа по общему правилу составляет 30 дней.
Важно помнить, что период для исполнения обязательства пользователем должен быть разумным. Иначе ответчик имеет шансы доказать невозможность выполнения требований. Если на первой стадии претензии собственника не удовлетворены, он направляет заявление в суд. Разбирательство возможно только в случае соблюдения досудебного порядка, поскольку истцу необходимы основания для расторжения договора и документальное подтверждение уведомления пользователя.
Судебная практика ст 619 гк рф
- Автострахование
- Жилищные споры
- Земельные споры
- Административное право
- Участие в долевом строительстве
- Семейные споры
- Гражданское право, ГК РФ
- Защита прав потребителей
- Трудовые споры, пенсии
- Главная
- Статья 619 ГК РФ. Досрочное расторжение договора по требованию арендодателя
Гражданский кодекс Российской Федерации:
Статья 619 ГК РФ.
По требованию арендодателя договор аренды может быть досрочно расторгнут судом в случаях, когда арендатор:
1) пользуется имуществом с существенным нарушением условий договора или назначения имущества либо с неоднократными нарушениями;
2) существенно ухудшает имущество;
3) более двух раз подряд по истечении установленного договором срока платежа не вносит арендную плату;
4) не производит капитального ремонта имущества в установленные договором аренды сроки, а при отсутствии их в договоре в разумные сроки в тех случаях, когда в соответствии с законом, иными правовыми актами или договором производство капитального ремонта является обязанностью арендатора.
Договором аренды могут быть установлены и другие основания досрочного расторжения договора по требованию арендодателя в соответствии с пунктом 2 статьи 450 настоящего Кодекса.
Арендодатель вправе требовать досрочного расторжения договора только после направления арендатору письменного предупреждения о необходимости исполнения им обязательства в разумный срок.
Вернуться к оглавлению документа: Гражданский кодекс РФ Часть 2 в действующей редакции
Комментарии к статье 619 ГК РФ, судебная практика применения
В п. 4 «Обзора судебной практики Верховного Суда РФ N 1 (2016)», утвержденного Президиумом Верховного Суда РФ 13.04.2016 года содержатся следующие разъяснения:
Стороны вправе предусмотреть в договоре аренды сумму компенсации, которая должна быть выплачена одной из сторон при немотивированном одностороннем отказе от исполнения договора.
Положения ст. 619, 620 ГК РФ не содержат запрета на досрочное расторжение договора аренды по иным основаниям, не предусмотренным в данных правовых нормах, в связи с чем стороны при заключении договора вправе установить основания для досрочного расторжения договора и условия такого расторжения. В частности, право на односторонний отказ от исполнения обязательства, связанного с осуществлением его сторонами предпринимательской деятельности, может быть обусловлено по соглашению сторон необходимостью выплаты определенной денежной суммы другой стороне обязательства.
См. подробнее об обстоятельствах дела в извлечении из «Обзора практики» во вложении ниже
В п. 23 Постановления Пленума ВАС РФ от 17.11.2011 N 73 «Об отдельных вопросах практики применения правил Гражданского кодекса Российской Федерации о договоре аренды» содержатся следующие разъяснения:
Досрочное расторжение договора аренды при невнесении арендной платы
Согласно статье 619 ГК РФ, если арендатор не вносит арендную плату более двух раз подряд по истечении установленного договором срока платежа, арендодатель вправе требовать досрочного расторжения договора аренды в судебном порядке.
Судам следует учитывать, что даже после уплаты долга арендодатель вправе в разумный срок предъявить иск о расторжении договора. Однако непредъявление такого требования в течение разумного срока с момента уплаты арендатором названного долга лишает арендодателя права требовать расторжения договора в связи с этим нарушением.
В пп. 5, 25-30 Информационного письма Президиума ВАС РФ от 11. 01.2002 N 66 «Обзор практики разрешения споров, связанных с арендой» содержатся следующие разъяснения:
01.2002 N 66 «Обзор практики разрешения споров, связанных с арендой» содержатся следующие разъяснения:
Договор аренды, заключенный (возобновленный) на неопределенный срок, может быть расторгнут по основаниям, предусмотренным статьей 619 ГК РФ.
..Вывод суда первой инстанции о том, что договор аренды, заключенный (возобновленный) на неопределенный срок, может быть прекращен только по основаниям, предусмотренным пунктом 2 статьи 610 ГК РФ, является несостоятельным, поскольку сам по себе факт заключения договора аренды на неопределенный срок не лишает арендодателя права на расторжение такого договора при существенном нарушении его условий арендатором.
Как видно из материалов дела, арендодатель на основании пунктов 1 и 3 части первой статьи 619 ГК РФ обратился в суд с иском о расторжении договора и выселении арендатора из занимаемого им помещения в связи с существенным нарушением последним условий договора, в том числе длительным невнесением арендной платы. В исковом заявлении арендодатель просил суд подтвердить факты нарушений, допущенных арендатором. До обращения в суд истцом были соблюдены требования части третьей статьи 619 и пункта 2 статьи 452 Кодекса, регламентирующие обязательные досудебные процедуры при расторжении договора.
В исковом заявлении арендодатель просил суд подтвердить факты нарушений, допущенных арендатором. До обращения в суд истцом были соблюдены требования части третьей статьи 619 и пункта 2 статьи 452 Кодекса, регламентирующие обязательные досудебные процедуры при расторжении договора.
Учитывая изложенное, суд апелляционной инстанции отметил, что арендодатель, определяя момент обращения в суд, правомерно руководствовался тридцатидневным сроком, установленным пунктом 2 статьи 452 ГК РФ, а не трехмесячным сроком, предусмотренным пунктом 2 статьи 610 Кодекса (п. 5 Информационного письма ВАС РФ № 66).
Основания досрочного расторжения договора аренды по требованию арендодателя, установленные в договоре в соответствии с частью второй статьи 619 ГК РФ, могут и не быть связаны с какими-либо нарушениями со стороны арендатора.
Часть первая статьи 619 ГК РФ предусматривает ряд оснований досрочного расторжения договора аренды по требованию арендодателя. Все эти основания касаются тех или иных нарушений договора со стороны арендатора.
Согласно части второй статьи 619 Кодекса договором аренды могут быть установлены и другие основания досрочного расторжения договора по требованию арендодателя в соответствии с пунктом 2 статьи 450 ГК РФ.
Однако то обстоятельство, что часть первая данной статьи в качестве оснований расторжения договора предусматривает исключительно нарушения условий договора со стороны арендатора, не означает, что другие основания расторжения, включаемые сторонами в договор, также должны быть связаны с нарушениями его условий.
Следует учитывать, что часть вторая статьи 619 ГК РФ отсылает ко всему пункту 2 статьи 450 Кодекса, а не только к подпункту 1 этого пункта. Поэтому стороны вправе оговорить любое основание досрочного расторжения договора в судебном порядке по требованию арендодателя, как связанное, так и не связанное с каким-либо нарушением (п. 25 Информационного письма ВАС РФ № 66).
В договоре аренды может быть предусмотрена возможность досрочного расторжения договора по требованию арендодателя в случае однократного невнесения арендатором арендной платы в установленный договором срок.
Истец в апелляционной жалобе настаивал на том, что спорное условие включено в договор в полном соответствии с требованиями части второй статьи 619 Кодекса. В силу пункта 3 части первой данной статьи основанием расторжения договора является невнесение арендной платы более двух раз подряд. Однократное нарушение арендатором установленных договором сроков внесения арендной платы есть другое основание расторжения договора аренды.
Суд апелляционной инстанции поддержал позицию истца, дополнительно отметив, что предусмотренное в части первой статьи 619 Кодекса основание включает в себя не только такой элемент, как «просрочка уплаты», но и еще два элемента: «более двух раз» и «подряд». С этой точки зрения в рассматриваемой ситуации стороны определили в договоре иное основание его расторжения, чем это предусмотрено пунктом 3 части первой статьи 619 ГК РФ (п. 26 Информационного письма ВАС РФ № 66).
В договоре аренды могут быть предусмотрены основания отказа арендодателя от исполнения договора и его расторжения во внесудебном порядке, в том числе связанные с нарушением арендатором того или иного условия договора (пункт 3 статьи 450 ГК РФ).
…В ходе судебного разбирательства установлено, что при заключении договора аренды по инициативе арендодателя в договор было включено условие, предусматривающее основание отказа арендодателя от исполнения договора в соответствии с пунктом 3 статьи 450 ГК РФ. В качестве такого основания было определено однократное невнесение арендатором арендной платы в установленный договором срок.
…Факт нарушения, с которым договор связывает право арендодателя отказаться от исполнения договора, доказан материалами дела и не оспаривается ответчиком. В силу пункта 3 статьи 450 Кодекса в этом случае договор считается расторгнутым, соответствующего решения суда не требуется.
Так как арендатор добровольно не освободил занимаемое нежилое помещение, арендодатель вправе был обратиться в суд с иском о его выселении в связи с прекращением договора аренды (п. 27 Информационного письма ВАС РФ № 66).
Договор аренды может быть расторгнут в связи с внесением арендной платы не в полном объеме, если данное нарушение суд признает существенным.
…Наличие статьи 619 ГК РФ, устанавливающей специальные правила досрочного расторжения договора, не исключает возможности досрочного расторжения договора аренды на основании статьи 450 Кодекса.
Согласно пункту 2 статьи 450 ГК РФ по требованию одной из сторон договор может быть расторгнут по решению суда при существенном нарушении договора другой стороной. Существенным признается нарушение договора одной из сторон, которое влечет для другой стороны такой ущерб, что она в значительной степени лишается того, на что была вправе рассчитывать при заключении договора.
Оценив обстоятельства дела и придя к выводу, что нарушение, допущенное ответчиком, является существенным, суд расторг договор аренды (п. 28 Информационного письма ВАС РФ № 66).
Если основанием расторжения договора аренды является неисполнение арендатором возложенных на него обязанностей, арендодатель до обращения в суд с иском о досрочном расторжении договора обязан направить арендатору письменное предупреждение о необходимости исполнения им обязательства в разумный срок (часть третья статьи 619 ГК РФ), а также предложение расторгнуть договор (пункт 2 статьи 452 Кодекса).
Как следовало из представленных истцом материалов, комитет направлял обществу такое предупреждение, в котором ставился вопрос о необходимости погашения задолженности по арендной плате.
Однако, по смыслу части третьей статьи 619 ГК РФ, указанное предупреждение само по себе не является предложением об изменении или расторжении договора аренды. Право требовать расторжения указанного договора возникает у арендодателя лишь в том случае, если в разумный срок арендатор не устранит соответствующие нарушения.
Так как часть третья статьи 619 Кодекса не регламентирует процедуру расторжения договора аренды, в отношениях по аренде действует общее правило, содержащееся в пункте 2 статьи 452 ГК РФ. Согласно этой норме требование об изменении или о расторжении договора может быть заявлено стороной в суд только после получения отказа другой стороны на предложение изменить или расторгнуть договор либо неполучения ответа в срок, указанный в предложении или установленный законом либо договором, а при его отсутствии — в тридцатидневный срок. (п. 29 Информационного письма ВАС РФ № 66).
(п. 29 Информационного письма ВАС РФ № 66).
Необходимым условием удовлетворения иска арендодателя о досрочном расторжении договора аренды на основании статьи 619 ГК РФ является установление в ходе судебного разбирательства факта получения арендатором письменного предупреждения арендодателя о необходимости исполнения договорного обязательства.
Согласно части третьей статьи 619 ГК РФ арендодатель вправе требовать досрочного расторжения договора только после направления арендатору письменного предупреждения о необходимости исполнения им обязательства в разумный срок.
Суд признал довод ответчика обоснованным, указав, что по смыслу части третьей статьи 619 Кодекса такое письменное предупреждение должно быть получено арендатором, чтобы он имел возможность исполнить договорное обязательство в разумный срок.
Как было установлено в ходе судебного разбирательства, истец со своей стороны не принял надлежащих мер, которые гарантировали бы получение ответчиком (арендатором) соответствующего письменного предупреждения (вручение под расписку, направление заказным письмом или с уведомлением о вручении) (п. 30 Информационного письма ВАС РФ № 66).
30 Информационного письма ВАС РФ № 66).
4. Стороны вправе предусмотреть в договоре аренды сумму компенсации, которая должна быть выплачена одной из сторон при немотивированном одностороннем отказе от исполнения договора.
Арендатор обратился в суд с иском к арендодателю о признании недействительным пункта договора, предусматривающего удержание арендодателем в полном объеме суммы задатка в качестве штрафа в случае досрочного расторжения арендатором договора аренды.
Суд первой инстанции, руководствуясь положениями п. 3 ст. 450, ст. 330 ГК РФ, удовлетворил заявленные требования в связи с тем, что установление в договоре штрафа за односторонний отказ от исполнения договора противоречит правовой природе неустойки как меры ответственности, применяемой за нарушение гражданских прав. Поскольку досрочное расторжение договора в одностороннем порядке не является гражданско-правовым нарушением (ненадлежащим исполнением обязательств), а обусловлено реализацией арендатором права, предоставленного договором, суды признали соответствующее условие договора противоречащим нормам гражданского законодательства.
Суд апелляционной инстанции и арбитражный суд округа поддержали выводы суда первой инстанции.
Судебная коллегия Верховного Суда Российской Федерации отменила судебные акты судов нижестоящих инстанций и отказала в удовлетворении иска по следующим основаниям.
Положения ст. 619, 620 ГК РФ не содержат запрета на досрочное расторжение договора аренды по иным основаниям, не предусмотренным в данных правовых нормах, в связи с чем стороны при заключении договора вправе установить основания для досрочного расторжения договора и условия такого расторжения. В частности, право на односторонний отказ от исполнения обязательства, связанного с осуществлением его сторонами предпринимательской деятельности, может быть обусловлено по соглашению сторон необходимостью выплаты определенной денежной суммы другой стороне обязательства.
Оспариваемым пунктом договора предусмотрена возможность расторжения договора до истечения срока его действия в одностороннем порядке по любым другим основаниям, кроме прямо указанных в договоре, однако условием такого расторжения является для арендатора — удержание арендодателем задатка, для арендодателя — уплата задатка в двойном размере.
В соответствии со ст. 329 ГК РФ исполнение обязательств может обеспечиваться неустойкой, залогом, удержанием имущества должника, поручительством, банковской гарантией, задатком и другими способами, предусмотренными законом или договором.
В данном случае задаток вносился и удерживался арендодателем в качестве гарантии надлежащего выполнения арендатором обязательств по договору, что не противоречит положениям указанной правовой нормы.
Таким образом, сторонами определен порядок расторжения договора, которым предусмотрено особое условие для досрочного немотивированного расторжения договора в одностороннем порядке, что не противоречит положениям ст. 329, 421 ГК РФ.
Поскольку удержание задатка арендодателем не связано с нарушением сторонами договорных обязательств, а при наличии определенных обстоятельств является условием для расторжения договора, квалификация судами суммы задатка в качестве неустойки основана на ошибочном толковании норм права.
В данном случае стороны при заключении договора определили сумму компенсации, которая должна быть выплачена одной из сторон при отказе от договора.
То, что такая компенсация поименована в договоре штрафом, не изменяет ее сути, которая состоит не в привлечении к ответственности стороны, решившей досрочно отказаться от договора, а, напротив, предоставляет возможность расторжения договора без объяснения причин любой из сторон.
Поскольку ГК РФ допускает любые предусмотренные законом или договором способы обеспечения обязательств, отсутствуют основания для признания недействительным оспариваемого пункта договора в части условия, предусматривающего право арендодателя удерживать задаток в качестве штрафа за досрочное расторжение договора, направленного на обеспечение исполнения арендатором условий договора и гарантирующего законные интересы арендодателя при его исполнении.
Статья 619 ГК РФ. Досрочное расторжение договора по требованию арендодателя (действующая редакция)
По требованию арендодателя договор аренды может быть досрочно расторгнут судом в случаях, когда арендатор:
1) пользуется имуществом с существенным нарушением условий договора или назначения имущества либо с неоднократными нарушениями;
2) существенно ухудшает имущество;
3) более двух раз подряд по истечении установленного договором срока платежа не вносит арендную плату;
4) не производит капитального ремонта имущества в установленные договором аренды сроки, а при отсутствии их в договоре в разумные сроки в тех случаях, когда в соответствии с законом, иными правовыми актами или договором производство капитального ремонта является обязанностью арендатора.
Договором аренды могут быть установлены и другие основания досрочного расторжения договора по требованию арендодателя в соответствии с пунктом 2 статьи 450 настоящего Кодекса.
Арендодатель вправе требовать досрочного расторжения договора только после направления арендатору письменного предупреждения о необходимости исполнения им обязательства в разумный срок.
- URL
- HTML
- BB-код
- Текст
Комментарий к ст. 619 ГК РФ
1. По общему правилу договор аренды расторгается по истечении срока его действия. Однако арендодатель может заявить требование о досрочном расторжении договора аренды в следующих случаях:
— использование арендатором имущества с существенным нарушением условий договора либо неоднократным нарушением;
— существенное ухудшение арендатором состояния имущества;
— невнесение более двух раз подряд в срок арендной платы;
— неосуществление капитального ремонта имущества при условии, что осуществление такого ремонта является обязанностью арендатора.
На практике возможно некоторое пересечение вышеприведенных оснований для досрочного расторжения договора аренды. Так, например, невнесение арендной платы арендатором является как специальным основанием для расторжения договора аренды, так и существенным нарушением условий договора либо неоднократным их нарушением. Из материалов судебной практики следует, что в этом случае суды анализируют период невнесения арендной платы и приходят к выводу, что, например, невнесение арендной платы в течение трех месяцев является общим основанием для расторжения договора аренды — существенное нарушение условий договора либо неоднократное их нарушение.
Также длительное невнесение арендной платы — общее основание расторжения договора аренды.
В договоре аренды могут быть установлены и иные основания для его расторжения по требованию арендодателя до истечения срока действия договора аренды.
Реализовать свое право на досрочное расторжение договора аренды при наличии основания арендодатель вправе только при условии направления арендатору письменного предупреждения о необходимости исполнения им обязательства в разумный срок.
2. Судебная практика:
— информационное письмо Президиума ВАС РФ от 11.01.2002 N 66;
— информационное письмо Президиума ВАС РФ от 05.05.1997 N 14;
— Постановление ФАС Северо-Западного округа от 10.10.2012 по делу N А56-59068/2011;
— Постановление Третьего арбитражного апелляционного суда от 03.03.2014 N 03АП-134/14;
— Постановление Пятого арбитражного апелляционного суда от 09.07.2014 N 05АП-6454/2014;
— Постановление Семнадцатого арбитражного апелляционного суда от 16.10.2013 N 17АП-9975/2013-АК;
— Постановление Пятнадцатого арбитражного апелляционного суда от 13.05.2013 N 15АП-4118/2013;
— Постановление ФАС Московского округа от 19.03.2007 N КГ-А40/1697-07-П-1,2.
Дело NПозиция ВС РФ.
Позиция ВС РФ: В договоре аренды, связанном с осуществлением его сторонами предпринимательской деятельности, может быть установлено, что компенсацией за одностороннее немотивированное расторжение является удержание арендодателем задатка или уплата им задатка в двойном размере
Применимые нормы: п. 3 ст. 310 , ст. ст. 329 , 381 , п. 4 ст. 421 , п. 2 ст . 450.1 , ст . ст . 619 , 620 ГК РФ
3 ст. 310 , ст. ст. 329 , 381 , п. 4 ст. 421 , п. 2 ст . 450.1 , ст . ст . 619 , 620 ГК РФ
Обзор судебной практики Верховного Суда Российской Федерации N 1 (утв. Президиумом Верховного Суда РФ 13.04.2016)
Право на односторонний отказ от исполнения обязательства, связанного с осуществлением его сторонами предпринимательской деятельности, по соглашению сторон может быть обусловлено выплатой определенной денежной суммы другой стороне обязательства.
Согласно условиям спорного договора арендодатель удерживал задаток, внесенный арендатором в качестве гарантии надлежащего выполнения обязательств по договору, что не противоречит положениям ст. 329 ГК РФ.
Стороны предусмотрели возможность досрочно расторгнуть договор в одностороннем порядке по любым основаниям, прямо не указанным в нем. Условием такого расторжения для арендатора являлось удержание арендодателем задатка, для арендодателя — уплата задатка в двойном размере.
Таким образом, стороны определили порядок расторжения договора, предусмотрев особое условие для его досрочного немотивированного расторжения в одностороннем порядке, что не противоречит положениям ст. ст. 329 , 421 ГК РФ. В данном случае при заключении договора была установлена сумма компенсации, которую должна выплатить одна из сторон при отказе от договора.
ст. 329 , 421 ГК РФ. В данном случае при заключении договора была установлена сумма компенсации, которую должна выплатить одна из сторон при отказе от договора.
Определение Верховного Суда РФ от 03.11.2015 N 305-ЭС15-6784 по делу N А40-53452/2014 (Судебная коллегия по экономическим спорам)
Все позиции высших судов к ст. 381 ГК РФ: см. Перечень позиций высших судов
Все позиции высших судов к ст. 421 ГК РФ: см. Перечень позиций высших судов
Все позиции высших судов к ст. 450.1 ГК РФ: см. Перечень позиций высших судов
Судебная практика по ст. 620 ГК РФ
- АПК РФ Статья 291.6. Рассмотрение кассационных жалобы, представления в Судебной коллегии Верховного Суда Российской Федерации
- АПК РФ Статья 71. Оценка доказательств
- ГК РФ Статья 310. Недопустимость одностороннего отказа от исполнения обязательства
- ГК РФ Статья 329. Способы обеспечения исполнения обязательств
- ГК РФ Статья 330. Понятие неустойки
- ГК РФ Статья 421.
 Свобода договора
Свобода договора - ГК РФ Статья 422. Договор и закон
- ГК РФ Статья 450. Основания изменения и расторжения договора
- ГК РФ Статья 620. Досрочное расторжение договора по требованию арендатора
- АПК РФ Статья 291.8. Определение об отказе в передаче кассационных жалобы, представления для рассмотрения в судебном заседании Судебной коллегии Верховного Суда Российской Федерации
- АПК РФ Статья 49. Изменение основания или предмета иска, изменение размера исковых требований, отказ от иска, признание иска, мировое соглашение
- ГК РФ Статья 309. Общие положения
 Свобода договора
Свобода договора- ГК РФ Статья 310. Недопустимость одностороннего отказа от исполнения обязательства
- ГК РФ Статья 450. Основания изменения и расторжения договора
- ГК РФ Статья 620. Досрочное расторжение договора по требованию арендатора
- ГК РФ Статья 668. Передача арендатору предмета договора финансовой аренды
- ГК РФ Статья 395. Ответственность за неисполнение денежного обязательства
- ГК РФ Статья 1107. Возмещение потерпевшему неполученных доходов
- АПК РФ Статья 291.6. Рассмотрение кассационных жалобы, представления в Судебной коллегии Верховного Суда Российской Федерации
- АПК РФ Статья 291.8. Определение об отказе в передаче кассационных жалобы, представления для рассмотрения в судебном заседании Судебной коллегии Верховного Суда Российской Федерации
- АПК РФ Статья 291.11. Основания для отмены или изменения Судебной коллегией Верховного Суда Российской Федерации судебных актов в порядке кассационного производства и присуждения компенсации за нарушение права на судопроизводство в разумный срок
- ГК РФ Статья 309. Общие положения
 Возмещение потерпевшему неполученных доходов
Возмещение потерпевшему неполученных доходов- ГК РФ Статья 310. Недопустимость одностороннего отказа от исполнения обязательства
- ГК РФ Статья 421. Свобода договора
- ГК РФ Статья 450. Основания изменения и расторжения договора
- ГК РФ Статья 619. Досрочное расторжение договора по требованию арендодателя
- ГК РФ Статья 620. Досрочное расторжение договора по требованию арендатора
- ГК РФ Статья 329. Способы обеспечения исполнения обязательств
- АПК РФ Статья 291.6. Рассмотрение кассационных жалобы, представления в Судебной коллегии Верховного Суда Российской Федерации
- АПК РФ Статья 291.8. Определение об отказе в передаче кассационных жалобы, представления для рассмотрения в судебном заседании Судебной коллегии Верховного Суда Российской Федерации
 Способы обеспечения исполнения обязательств
Способы обеспечения исполнения обязательств- АПК РФ Статья 291.6. Рассмотрение кассационных жалобы, представления в Судебной коллегии Верховного Суда Российской Федерации
- ГК РФ Статья 310. Недопустимость одностороннего отказа от исполнения обязательства
- ГК РФ Статья 431. Толкование договора
- ГК РФ Статья 450. Основания изменения и расторжения договора
- ГК РФ Статья 620. Досрочное расторжение договора по требованию арендатора
- АПК РФ Статья 291.8. Определение об отказе в передаче кассационных жалобы, представления для рассмотрения в судебном заседании Судебной коллегии Верховного Суда Российской Федерации
- ГК РФ Статья 310. Недопустимость одностороннего отказа от исполнения обязательства
- ГК РФ Статья 606. Договор аренды
- ГК РФ Статья 614. Арендная плата
- ГК РФ Статья 619. Досрочное расторжение договора по требованию арендодателя
- ГК РФ Статья 620. Досрочное расторжение договора по требованию арендатора
- ГК РФ Статья 329. Способы обеспечения исполнения обязательств
- ГК РФ Статья 421. Свобода договора
- АПК РФ Статья 291.6. Рассмотрение кассационных жалобы, представления в Судебной коллегии Верховного Суда Российской Федерации
- АПК РФ Статья 71. Оценка доказательств
- ГК РФ Статья 309. Общие положения
 Недопустимость одностороннего отказа от исполнения обязательства
Недопустимость одностороннего отказа от исполнения обязательства- ГК РФ Статья 450. Основания изменения и расторжения договора
- ГК РФ Статья 620. Досрочное расторжение договора по требованию арендатора
- АПК РФ Статья 291.6. Рассмотрение кассационных жалобы, представления в Судебной коллегии Верховного Суда Российской Федерации
- АПК РФ Статья 71. Оценка доказательств
- ГК РФ Статья 450. Основания изменения и расторжения договора
- ГК РФ Статья 619. Досрочное расторжение договора по требованию арендодателя
- ГК РФ Статья 620. Досрочное расторжение договора по требованию арендатора
- ЗК РФ Статья 45. Основания прекращения права постоянного (бессрочного) пользования земельным участком, права пожизненного наследуемого владения земельным участком
- ЗК РФ Статья 46. Основания прекращения аренды земельного участка
- ГК РФ Статья 451. Изменение и расторжение договора в связи с существенным изменением обстоятельств
- АПК РФ Статья 291.11. Основания для отмены или изменения Судебной коллегией Верховного Суда Российской Федерации судебных актов в порядке кассационного производства и присуждения компенсации за нарушение права на судопроизводство в разумный срок
- АПК РФ Статья 291.6. Рассмотрение кассационных жалобы, представления в Судебной коллегии Верховного Суда Российской Федерации
 Основания изменения и расторжения договора
Основания изменения и расторжения договора- АПК РФ Статья 125. Форма и содержание искового заявления
- АПК РФ Статья 126. Документы, прилагаемые к исковому заявлению
- АПК РФ Статья 291.6. Рассмотрение кассационных жалобы, представления в Судебной коллегии Верховного Суда Российской Федерации
- АПК РФ Статья 291.8. Определение об отказе в передаче кассационных жалобы, представления для рассмотрения в судебном заседании Судебной коллегии Верховного Суда Российской Федерации
- АПК РФ Статья 291.11. Основания для отмены или изменения Судебной коллегией Верховного Суда Российской Федерации судебных актов в порядке кассационного производства и присуждения компенсации за нарушение права на судопроизводство в разумный срок
- ГК РФ Статья 620. Досрочное расторжение договора по требованию арендатора
 Документы, прилагаемые к исковому заявлению
Документы, прилагаемые к исковому заявлению- ГК РФ Статья 431. Толкование договора
- ГК РФ Статья 450. Основания изменения и расторжения договора
- ГК РФ Статья 620. Досрочное расторжение договора по требованию арендатора
- АПК РФ Статья 291.6. Рассмотрение кассационных жалобы, представления в Судебной коллегии Верховного Суда Российской Федерации
- АПК РФ Статья 291. 9. Определение о передаче кассационных жалобы, представления вместе с делом для рассмотрения в судебном заседании Судебной коллегии Верховного Суда Российской Федерации
- АПК РФ Статья 291.11. Основания для отмены или изменения Судебной коллегией Верховного Суда Российской Федерации судебных актов в порядке кассационного производства и присуждения компенсации за нарушение права на судопроизводство в разумный срок
 9. Определение о передаче кассационных жалобы, представления вместе с делом для рассмотрения в судебном заседании Судебной коллегии Верховного Суда Российской Федерации
9. Определение о передаче кассационных жалобы, представления вместе с делом для рассмотрения в судебном заседании Судебной коллегии Верховного Суда Российской Федерации- ГК РФ Статья 12. Способы защиты гражданских прав
- ГК РФ Статья 15. Возмещение убытков
- ГК РФ Статья 310. Недопустимость одностороннего отказа от исполнения обязательства
- ГК РФ Статья 393. Обязанность должника возместить убытки
- ГК РФ Статья 431. Толкование договора
- ГК РФ Статья 450. Основания изменения и расторжения договора
- ГК РФ Статья 606. Договор аренды
- ГК РФ Статья 611. Предоставление имущества арендатору
- ГК РФ Статья 612. Ответственность арендодателя за недостатки сданного в аренду имущества
- ГК РФ Статья 620. Досрочное расторжение договора по требованию арендатора
- ГК РФ Статья 1064. Общие основания ответственности за причинение вреда
- ГК РФ Статья 1082. Способы возмещения вреда
- АПК РФ Статья 291.6. Рассмотрение кассационных жалобы, представления в Судебной коллегии Верховного Суда Российской Федерации
- АПК РФ Статья 291.8. Определение об отказе в передаче кассационных жалобы, представления для рассмотрения в судебном заседании Судебной коллегии Верховного Суда Российской Федерации
- ГК РФ Статья 309. Общие положения
 Досрочное расторжение договора по требованию арендатора
Досрочное расторжение договора по требованию арендатораДоговор-Юрист
— это юристы, кодексы и бланки
Кодексы РФ
Типовые договоры
Активные юристы
Лучшие юристы
Обновления кодексов
Ответы юристов
Судебная практика по ст. 620 ГК РФ
Раздел посвящён судебной практике Российской Федерации. Здесь вы найдёте судебные решения Верховного Суда Российской Федерации. База судебной прпктики Договор-Юрист.Ру ежедневно проверяется и обновляется.
База судебной прпктики Договор-Юрист.Ру ежедневно проверяется и обновляется.
На сайте предусмотрен удобный поиск судебной практики по статьям, например «ст. 620 ГК РФ», вы увидите все свежие судебные решения с упоминанием ст. 620 ГК РФ.
Копирование материалов с сайта «Договор-Юрист. Ру» возможно только с разрешения администрации сайта и с индексируемой ссылкой на источник.
Под «бесплатными юридическими консультациями» подразумеваются ответы на типовые вопросы, справочная информация по статьям кодексов и законов
Другие статьи:
- За сколько дней открывать спор на али Продавец выслал товар После того, как продавец вышлет ваш товар и вы получите заветный трек номер, в вашем заказе появится таймер защиты. Это, пожалуй, самый важный таймер, на который стоит ориентироваться. Пока тикает таймер – деньги находятся на счету у […]
- Проскурин геннадий алексеевич адвокат Коллегия адвокатов «Правовая Защита»
ВНИМАНИЕ. Список адвокатов формируется по фамилиям со второй буквы по алфавиту, но в обратном порядке! Проще смотреть снизу вверх.
Все адвокаты указаны с номером, который им присвоен в Адвокатской Палате Липецкой области. […]
- Офицер это льготы Льготы военнослужащим в 2019 году Военнослужащие — это одна из категорий граждан, которая напрямую зависит от того, что предлагает и предоставляет государство. В 2017 году произошло внесение нескольких изменений, которые коснулись именно тех, кто проходит […]
- Аренда квартиры единый налог Налог на аренду: как сэкономить рантье? Prosto страх берет, когда почитаешь про рейды налоговиков, «прижимающих к стенке» арендодателей, не платящих со своих доходов налоги. Что же делать гражданам, сдающим жилье, чтобы остаться в рамках закона и не лишиться […]
- Экспертиза в петрозаводске Центры экспертиз в Петрозаводске
1. Эксперт-Cервис-Карелия на ул. Кирова, 5
Адрес: Петрозаводск, ул. Кирова, 5, оф. 411 Телефон: +7 (8142) 26-00-58; +7 (921) 726-00-58 Режим работы: пн-пт 09:00-19:30
2. АНО центр Бухгалтерских Экспертиз на просп. Карельский, […]
- Приказ установить нормы списания Приказ о нормах расхода топлива и ГСМ Приказ о нормах расхода топлива и ГСМ – документ, применяющийся для определения норм расхода горюче-смазочных материалов, учитывая при этом марку, модификацию автомобиля, а так же выполняемые работы конкретным транспортным […]
- Заявление на проведение аккредитации Вопросы аккредитации СВР России осуществляет аккредитацию органов по сертификации и испытательных лабораторий (центров), выполняющих работы по оценке (подтверждению) соответствия в отношении продукции (работ, услуг), используемой в целях защиты сведений, […]
 Список адвокатов формируется по фамилиям со второй буквы по алфавиту, но в обратном порядке! Проще смотреть снизу вверх.
Все адвокаты указаны с номером, который им присвоен в Адвокатской Палате Липецкой области. […]
Список адвокатов формируется по фамилиям со второй буквы по алфавиту, но в обратном порядке! Проще смотреть снизу вверх.
Все адвокаты указаны с номером, который им присвоен в Адвокатской Палате Липецкой области. […] Эксперт-Cервис-Карелия на ул. Кирова, 5
Адрес: Петрозаводск, ул. Кирова, 5, оф. 411 Телефон: +7 (8142) 26-00-58; +7 (921) 726-00-58 Режим работы: пн-пт 09:00-19:30
2. АНО центр Бухгалтерских Экспертиз на просп. Карельский, […]
Эксперт-Cервис-Карелия на ул. Кирова, 5
Адрес: Петрозаводск, ул. Кирова, 5, оф. 411 Телефон: +7 (8142) 26-00-58; +7 (921) 726-00-58 Режим работы: пн-пт 09:00-19:30
2. АНО центр Бухгалтерских Экспертиз на просп. Карельский, […]Статья 619.
 Досрочное расторжение договора по требованию арендодателя
Досрочное расторжение договора по требованию арендодателяПо требованию арендодателя договор аренды может быть досрочно расторгнут судом в случаях, когда арендатор:
1) пользуется имуществом с существенным нарушением условий договора или назначения имущества либо с неоднократными нарушениями;
2) существенно ухудшает имущество;
3) более двух раз подряд по истечении установленного договором срока платежа не вносит арендную плату;
4) не производит капитального ремонта имущества в установленные договором аренды сроки, а при отсутствии их в договоре в разумные сроки в тех случаях, когда в соответствии с законом, иными правовыми актами или договором производство капитального ремонта является обязанностью арендатора.
Договором аренды могут быть установлены и другие основания досрочного расторжения договора по требованию арендодателя в соответствии с пунктом 2 статьи 450 настоящего Кодекса.
* Арендодатель вправе требовать досрочного расторжения договора только после направления арендатору письменного предупреждения о необходимости исполнения им обязательства в разумный срок.
Комментарий к статье 619 Гражданского кодекса РФ
на основе судебной практики
Постановление Пленума ВАС РФ от 17.11.2011 N 73 «Об отдельных вопросах практики применения правил Гражданского кодекса Российской Федерации о договоре аренды»
Пункт 23. Согласно статье 619 ГК РФ, если арендатор не вносит арендную плату более двух раз подряд по истечении установленного договором срока платежа, арендодатель вправе требовать досрочного расторжения договора аренды в судебном порядке.
Судам следует учитывать, что даже после уплаты долга арендодатель вправе в разумный срок предъявить иск о расторжении договора. Однако непредъявление такого требования в течение разумного срока с момента уплаты арендатором названного долга лишает арендодателя права требовать расторжения договора в связи с этим нарушением. (п. 23 введен Постановлением Пленума ВАС РФ от 25.01.2013 N 13)
Комментарий к пункту 23 Постановления Пленума ВАС РФ от 17.11.2011 N 7323
Разъяснения, содержащиеся в пункте 23 Постановления N 73, которые приводит Комитет в обоснование своей жалобы, касаются права арендодателя предъявить иск о расторжении договора в разумный срок после уплаты долга и не содержат безусловной обязанности суда удовлетворить такое требование.
Отказ арендодателя от исполнения Договора, чтобы считаться правомерным, должен быть произведен в период имеющейся (продолжающейся) просрочки арендатора.
Определение ВС РФ от 3 ноября 2015 г. N 305-ЭС15-6784
Поскольку положения статей 619, 620 Гражданского кодекса не содержат запрета на досрочное расторжение договора аренды по иным основаниям, не предусмотренным в данных правовых нормах, стороны при заключении договора вправе установить основания для досрочного расторжения договора и условия такого расторжения. В частности, право на односторонний отказ от исполнения обязательства, связанного с осуществлением его сторонами предпринимательской деятельности, может быть обусловлено по соглашению сторон необходимостью выплаты определенной денежной суммы другой стороне обязательства.
То, что такая компенсация поименована в договоре штрафом, не изменяет ее сути, которая состоит не в привлечении к ответственности стороны, решившей досрочно отказаться от договора, а напротив, предоставляет возможность расторжения договора без объяснения причин любой из сторон.
Постановление Арбитражного суда Западно-Сибирского округа от 21.09.2016 N Ф04-3353/2016 по делу N А03-21693/2015.
официальный сайт компании Herbalife Nutrition в России
Узнайте, находится ли Ваш вес в пределах нормы или нет, и получите персональные рекомендации
Ваш вес ниже нормы
индекс массы тела —
Узнайте как питаться сбалансированно и увеличить вес до уровня нормы, для этого заполните форму заявки чтобы связаться с Консультантом по здоровому образу жизни в Вашем городе. При изменении рациона питания рекомендуем Вам консультироваться с лечащим врачом.
Ваш вес в пределах нормы
индекс массы тела —
Чтобы поддерживать хорошее самочувствие и вес в пределах нормы, обратитесь к Консультанту по здоровому образу жизни в Вашем городе, заполнив форму заявки. Консультант познакомит Вас с принципами сбалансированного питания, научит завтракать и ужинать с пользой для организма.
Консультант познакомит Вас с принципами сбалансированного питания, научит завтракать и ужинать с пользой для организма.
Ваш вес немного превышает нормальный
индекс массы тела —
Что может стать причиной плохого самочувствия и фактором риска возникновения заболеваний. Чтобы узнать как снизить вес и улучшить самочувствие, заполните форму заявки для связи с Консультантом по здоровому образу жизни в Вашем городе. При изменении рациона питания рекомендуем Вам консультироваться с лечащим врачом.
У Вас избыточный вес
индекс массы тела —
Который может стать причиной плохого самочувствия и фактором риска возникновения заболеваний. Чтобы узнать как снизить вес и улучшить самочувствие, заполните форму заявки для связи с Консультантом по здоровому образу жизни в Вашем городе. При изменении рациона питания рекомендуем Вам консультироваться слечащим врачом.
Чтобы узнать как снизить вес и улучшить самочувствие, заполните форму заявки для связи с Консультантом по здоровому образу жизни в Вашем городе. При изменении рациона питания рекомендуем Вам консультироваться слечащим врачом.
Взрослые мужчины, пережившие сексуальное насилие в детстве: распространенность, характеристики сексуального насилия и долгосрочные последствия
Сексуальное насилие в детстве (CSA) было определено как мощный фактор риска сексуальной дисфункции. Определенные характеристики опыта жестокого обращения, такие как повторяющееся насилие, по-видимому, влияют на риск развития сексуальной дисфункции. Несмотря на убедительные данные о том, что CSA может быть вредным для сексуальной функции, нет единого мнения о точных механизмах, которые приводят к этим трудностям.
Обобщить самые современные исследования связи между CSA и сексуальной функцией женщин.
Был проведен обзор опубликованной литературы, посвященной распространенности сексуальной дисфункции среди женщин с СЭД в анамнезе, различных типов сексуальных дисфункций и механизмов, предложенных для объяснения связи между СЭД и более поздними сексуальными трудностями.
Обзор рецензируемой литературы.
Женщины, подвергшиеся насилию в прошлом, сообщают о более высоких показателях сексуальной дисфункции по сравнению с их сверстницами, не подвергавшимися насилию.Сексуальные проблемы, о которых чаще всего сообщают женщины, подвергшиеся жестокому обращению, включают проблемы с сексуальным желанием и сексуальным возбуждением. Механизмы, которые были предложены для объяснения связи между CSA и сексуальной дисфункцией, включают когнитивные ассоциации с сексуальностью, сексуальные схемы себя, активацию симпатической нервной системы, образ тела и уважение, а также стыд и вину.
Женщины с CSA в анамнезе представляют уникальную группу населения в литературе по сексуальному здоровью. Обзор механизмов, предложенных для объяснения взаимосвязи между CSA и сексуальным здоровьем, показывает, что отсутствие положительных эмоций, связанных с сексуальностью, а не усиление отрицательных эмоций, по-видимому, более важно для сексуального здоровья женщин с CSA в анамнезе.Исследования в области лечения показали, что секс-терапия, основанная на осознанности, и экспрессивное письмо особенно эффективны для этой группы. Необходимы дальнейшие исследования, чтобы прояснить механизмы, которые приводят к сексуальной дисфункции у женщин, подвергшихся насилию в прошлом, чтобы обеспечить более целенаправленное лечение сексуальной дисфункции у женщин, подвергшихся насилию в прошлом.
Пулверман К.С., Килимник К.Д., Местон К.М. Влияние сексуального насилия в детстве на сексуальное здоровье женщин: всесторонний обзор. Sex Med Rev 2018; 6: 188–200.
Укорочения тайтина, вызывающие дилатационную кардиомиопатию
Субъекты исследования
Идиопатическая дилатационная кардиомиопатия была диагностирована у 312 субъектов из трех независимых групп: группы А, группы В и группы С (таблицы 9, 10 и 11 соответственно, в дополнительном приложении). Гипертрофическая кардиомиопатия была диагностирована у 231 человека; 249 человек без известной кардиомиопатии служили контролем. Сводные характеристики каждой группы и когорты представлены в таблице 3 в дополнительном приложении.
Гипертрофическая кардиомиопатия была диагностирована у 231 человека; 249 человек без известной кардиомиопатии служили контролем. Сводные характеристики каждой группы и когорты представлены в таблице 3 в дополнительном приложении.
Используя геномную ДНК, выделенную от субъектов с дилатационной кардиомиопатией в группах A и B, а также от субъектов с гипертрофической кардиомиопатией и контрольной группы, мы захватили и провели секвенирование следующего поколения 145 т.п.н. гена TTN , включая все аннотированные экзоны и сайты сплайсинга. Для каждого субъекта более 97% целевых оснований секвенировали не менее 20 раз (данные не показаны).Из геномной ДНК, выделенной от пациентов с дилатационной кардиомиопатией, мы определили последовательностей TTN с помощью традиционного дидезокси-секвенирования.
После исключения вариантов TTN с частотой 0,01 или выше в рамках проекта «1000 геномов» (www.1000genomes.org/data) или присутствующих у всех 792 субъектов исследования, мы выявили 951 редкий миссенс (частота минорного аллеля <0,01), бессмыслица , сдвиг рамки считывания или сплайсинг или номер копии вариантов TTN , которые, по прогнозам, изменят аминокислотную последовательность титина (таблицы 4 и 5 в дополнительном приложении).У каждого субъекта исследования был примерно 1 редкий вариант миссенс (диапазон средних значений на группу или когорту от 0,91 до 1,45 на субъекта).
Поскольку считается, что транскрипт TTN novex-3 (рис. 1) не взаимодействует с M-полосой саркомера, а его экспрессия в сердце составляет примерно одну двадцатую от полноразмерных изоформ тайтина (неопубликованные данные), мы не изучали варианты, которые изменяют исключительно транскрипты novex-3. Мы выделили для анализа четыре типа вариантов — i.т. е., нонсенс, сдвиг рамки считывания, сплайсинг и число копий, которые, как предполагается, (в отличие от большинства миссенс-вариантов) оказывают сильное влияние на структуру полноразмерных полипептидов тайтина (рис. 1). Мы классифицируем их как варианты усечения ТТН .
1). Мы классифицируем их как варианты усечения ТТН .
 34). Субъекты с дилатационной кардиомиопатией
34). Субъекты с дилатационной кардиомиопатией Бессмысленные варианты и варианты со сдвигом рамки
Мы выявили 44 уникальных бессмысленных варианта или варианта со сдвигом рамки, которые изменяют полноразмерный тайтин у 47 субъектов с дилатационной кардиомиопатией (таблица 1 и таблица 6 в дополнительном приложении). В когортах и группах, исследованных с помощью секвенирования нового поколения, доля лиц с этими вариантами была выше среди лиц с дилатационной кардиомиопатией в группах А и В (21 из 92 человек [23%] и 14 из 71 человека [20%]. ] соответственно), чем среди субъектов с гипертрофической кардиомиопатией (2 из 231 [1%], P=2×10 -12 ) или контрольных субъектов (2 из 249 [1%], P=3×10 — 13 ).Такие варианты были у 12 из 149 человек (8%) с дилатационной кардиомиопатией в группе С, обследованных с помощью традиционного дидезокси-секвенирования. Мы наблюдали сильную косегрегацию (значение LOD, 9,3) нонсенс-вариантов и вариантов со сдвигом рамки с клиническим статусом среди 60 членов 16 семей, страдающих дилатационной кардиомиопатией (рис. 2, а также таблицы 12 и таблицы 13 в дополнительном приложении), что указывает на вероятность приблизительно 1 в 10 9 , что выделение этих вариантов ТТН произошло случайно.У одного субъекта с дилатационной кардиомиопатией в возрасте 17 лет был один редкий вариант на каждом аллеле TTN : ранее описанная патогенная миссенс-мутация 9 и нонсенс-вариант, унаследованный от ее матери (рис. 2 в дополнительном приложении). Во всех других семьях имело место совместное наследование дилатационной кардиомиопатии и одного варианта со сдвигом рамки TTN или нонсенс-варианта. Среди 32 членов семьи старше 40 лет, по которым имелись данные, пенетрантность укороченных мутаций TTN составила более 95%.
Мы наблюдали сильную косегрегацию (значение LOD, 9,3) нонсенс-вариантов и вариантов со сдвигом рамки с клиническим статусом среди 60 членов 16 семей, страдающих дилатационной кардиомиопатией (рис. 2, а также таблицы 12 и таблицы 13 в дополнительном приложении), что указывает на вероятность приблизительно 1 в 10 9 , что выделение этих вариантов ТТН произошло случайно.У одного субъекта с дилатационной кардиомиопатией в возрасте 17 лет был один редкий вариант на каждом аллеле TTN : ранее описанная патогенная миссенс-мутация 9 и нонсенс-вариант, унаследованный от ее матери (рис. 2 в дополнительном приложении). Во всех других семьях имело место совместное наследование дилатационной кардиомиопатии и одного варианта со сдвигом рамки TTN или нонсенс-варианта. Среди 32 членов семьи старше 40 лет, по которым имелись данные, пенетрантность укороченных мутаций TTN составила более 95%.
Варианты сплайсинга
Мы идентифицировали 17 вариантов TTN у 19 субъектов с дилатационной кардиомиопатией, которые, по прогнозам, изменяют сплайсинг РНК, в том числе 11, которые изменяют абсолютно консервативные нуклеотиды сайта сплайсинга (таблица 1 и таблица 7 в дополнительном приложении). Секвенирование РНК образцов сердечной ткани двух субъектов с вариантами сплайсинга (один взрослый ребенок от кровнородственного брака с гомозиготным вариантом сплайсинга [данные не показаны], другой описан на рис.3 в дополнительном приложении) подтвердил аномальный сплайсинг TTN . Среди субъектов, исследованных с использованием секвенирования нового поколения, мы наблюдали значительное обогащение вариантов сплайсинга у субъектов с дилатационной кардиомиопатией в группе А (с 15 из 92 субъектов [16%] пораженных) и группе В (с 3 из 71 субъекта [16]. 4%] поражены) по сравнению с субъектами с гипертрофической кардиомиопатией (только у 1 из 231 субъекта [<1%] поражены, P = 7 × 10 -8 ) и контрольными субъектами (с 5 из 249 субъектов [2%]). ] затронуты, P=9×10 -6 ).Мы обнаружили полную косегрегацию вариантов сплайсинга и дилатационную кардиомиопатию (показатель lod, 1,8, что указывает на связь, которая была в 60 раз более вероятной, чем случайная) среди 11 членов трех семей (рис.
Секвенирование РНК образцов сердечной ткани двух субъектов с вариантами сплайсинга (один взрослый ребенок от кровнородственного брака с гомозиготным вариантом сплайсинга [данные не показаны], другой описан на рис.3 в дополнительном приложении) подтвердил аномальный сплайсинг TTN . Среди субъектов, исследованных с использованием секвенирования нового поколения, мы наблюдали значительное обогащение вариантов сплайсинга у субъектов с дилатационной кардиомиопатией в группе А (с 15 из 92 субъектов [16%] пораженных) и группе В (с 3 из 71 субъекта [16]. 4%] поражены) по сравнению с субъектами с гипертрофической кардиомиопатией (только у 1 из 231 субъекта [<1%] поражены, P = 7 × 10 -8 ) и контрольными субъектами (с 5 из 249 субъектов [2%]). ] затронуты, P=9×10 -6 ).Мы обнаружили полную косегрегацию вариантов сплайсинга и дилатационную кардиомиопатию (показатель lod, 1,8, что указывает на связь, которая была в 60 раз более вероятной, чем случайная) среди 11 членов трех семей (рис. 2 и таблица 13 в дополнительном приложении).
2 и таблица 13 в дополнительном приложении).
Варианты номеров копий
Мы оценили количество копий в TTN у субъектов с дилатационной кардиомиопатией в группах A и B путем сравнения распределения прочтений последовательностей среди субъектов (рис. 4 в дополнительном приложении).Традиционное дидезокси-секвенирование, используемое для образцов от субъектов с дилатационной кардиомиопатией в группе С, не позволяло проводить эти анализы. Один вариант числа копий был идентифицирован у субъекта с дилатационной кардиомиопатией. Было предсказано, что эта тандемная вставка размером 28 т.п.н. (охватывающая интроны с 71 по 124) включает 13% внутреннего дублирования части тайтина; его наличие было подтверждено у пробанда и отсутствовало у двух здоровых родственников (рис. 5 в дополнительном приложении). Никаких изменений числа копий TTN не наблюдалось среди субъектов с гипертрофической кардиомиопатией или контрольных субъектов (всего 480).
Последствия
TTN Усеченных вариацийПоскольку TTN бессмысленный вариант, варианты со сдвигом рамки считывания, сплайсингом и числом копий, которые, по прогнозам, существенно изменяют структуру тайтина, значительно чаще встречались среди субъектов с дилатационной кардиомиопатией, чем среди субъектов с гипертрофической кардиомиопатией ( P=3×10 -16 ) или в контроле (P=9×10 -14 ) и наследовались совместно с дилатационной кардиомиопатией в семьях (комбинированный показатель lod, 11.1), мы пришли к выводу, что эти варианты вызывают дилатационную кардиомиопатию. Шесть мутаций TTN присутствовали у двух субъектов; результаты анализов одной пары субъектов соответствовали общему гаплотипу (данные не показаны).
Чтобы консервативно оценить частоту усеченных мутаций TTN у субъектов с дилатационной кардиомиопатией, мы увеличили нашу популяцию субъектов с дилатационной кардиомиопатией в группе A, добавив 40 субъектов, которые были набраны одновременно с другими субъектами в группе и у которых TTN последовательностей не были проанализированы, поскольку исследования выявили патогенную мутацию в другом гене дилатационной кардиомиопатии. После этого добавления частоты усеченных мутаций TTN среди субъектов с дилатационной кардиомиопатией в группах A и B составили 28% и 24% соответственно (P = 0,74). У субъектов группы С, образцы которых исследовались с использованием другой платформы секвенирования ДНК, было значительно меньше мутаций TTN (9%, P<0,001). Частота мутаций TTN существенно не отличалась между субъектами с дилатационной кардиомиопатией и без нее в семейном анамнезе (P = 0.36) (таблица 3 в дополнительном приложении).
После этого добавления частоты усеченных мутаций TTN среди субъектов с дилатационной кардиомиопатией в группах A и B составили 28% и 24% соответственно (P = 0,74). У субъектов группы С, образцы которых исследовались с использованием другой платформы секвенирования ДНК, было значительно меньше мутаций TTN (9%, P<0,001). Частота мутаций TTN существенно не отличалась между субъектами с дилатационной кардиомиопатией и без нее в семейном анамнезе (P = 0.36) (таблица 3 в дополнительном приложении).
Усеченные мутации TTN , обнаруженные у субъектов с дилатационной кардиомиопатией, были неслучайно распределены в тайтине (рис. 1): они были чрезмерно представлены в области полосы A по сравнению с остатком N2BA (P = 0,0004) или N2B (P = 0,01) и отсутствовали в областях Z-диска и М-полосы (P=0,006 и P=0,001 для сравнения с N2BA и с N2B соответственно). Пространственное распределение 10 вариантов, обнаруженных у субъектов с гипертрофической кардиомиопатией и контрольных субъектов (рис. 1), отличалось от распределения вариантов, обнаруженных у субъектов с дилатационной кардиомиопатией (P = 0.001). Варианты у субъектов с гипертрофической кардиомиопатией и контрольных субъектов были менее обогащены областью A-диапазона тайтина (40% против 84% для мутаций у субъектов с дилатационной кардиомиопатией, P = 0,006) и включали варианты Z-диапазона ( 20% против 0%, р=0,01).
1), отличалось от распределения вариантов, обнаруженных у субъектов с дилатационной кардиомиопатией (P = 0.001). Варианты у субъектов с гипертрофической кардиомиопатией и контрольных субъектов были менее обогащены областью A-диапазона тайтина (40% против 84% для мутаций у субъектов с дилатационной кардиомиопатией, P = 0,006) и включали варианты Z-диапазона ( 20% против 0%, р=0,01).
Клинические характеристики
Таблица 2. Таблица 2. Характеристики субъектов с дилатационной кардиомиопатией в зависимости от наличия или отсутствия мутаций TTN . Рисунок 2. Рисунок 2. Оценка Каплана-Мейера возраста на момент постановки диагноза и клинического прогрессирования у субъектов с дилатационной кардиомиопатией, вызванной мутациями TTN . Панель A показывает данные на момент постановки диагноза, а панели B, C и D показывают отсутствие трансплантации сердца, имплантации вспомогательного устройства для левого желудочка (LVAD) и смерти от сердечных причин. Панели A и B показывают данные для 67 субъектов, у которых были мутации, и 228, у которых их не было. На панелях C и D показаны данные для 94 субъектов (из 19 семей), у которых были мутации усечения (рис. 2 в дополнительном приложении; также см. дополнительное приложение для описания влияния пола).Заштрихованные метки обозначают субъекты с подвергнутыми цензуре данными.
Панели A и B показывают данные для 67 субъектов, у которых были мутации, и 228, у которых их не было. На панелях C и D показаны данные для 94 субъектов (из 19 семей), у которых были мутации усечения (рис. 2 в дополнительном приложении; также см. дополнительное приложение для описания влияния пола).Заштрихованные метки обозначают субъекты с подвергнутыми цензуре данными.
Не было выявлено существенных различий между субъектами с усеченными мутациями TTN и без них в отношении возраста на момент постановки диагноза, конечно-диастолических размеров левого желудочка, фракции выброса или частоты трансплантации сердца, имплантации вспомогательного устройства для левого желудочка, и смерть от сердечных причин (таблица 2 и рисунок 2A и 2B).
Разнообразие клинических проявлений дилатационной кардиомиопатии отражалось у субъектов в группах А, В и С.Субъекты в группе А были более склонны к семейному заболеванию, чем субъекты в группе В (P = 6 × 10 -13 ) или группе C (P = 0,005). Субъекты в группе B (у которых клинические признаки были установлены во время оценки трансплантации сердца) чаще имели тяжелую дилатационную кардиомиопатию, чем субъекты в группах A и C, со значительно увеличенным конечно-диастолическим диаметром левого желудочка (P = 1 × 10 -9). и P=3×10 -6 соответственно) и снижение фракции выброса левого желудочка (P=0.03 и P = 0,0001 соответственно) (таблица 2 и рис. 1 в дополнительном приложении).
Субъекты в группе B (у которых клинические признаки были установлены во время оценки трансплантации сердца) чаще имели тяжелую дилатационную кардиомиопатию, чем субъекты в группах A и C, со значительно увеличенным конечно-диастолическим диаметром левого желудочка (P = 1 × 10 -9). и P=3×10 -6 соответственно) и снижение фракции выброса левого желудочка (P=0.03 и P = 0,0001 соответственно) (таблица 2 и рис. 1 в дополнительном приложении).
Частота трансплантации сердца, имплантации вспомогательного устройства для левого желудочка и смерти от сердечных причин среди субъектов и членов их семей с мутациями TTN , по-видимому, не зависит от типа мутации, но зависит от пола субъектов (рис. 2С и 2D). Средний (±SD) возраст на момент этих нежелательных явлений у 94 носителей мутации в 19 семьях составлял 68±5 лет для 33 женщин и 56±3 года для 61 мужчины (P=4×10 -5 ). (Инжир.2 в дополнительном приложении).
Рисунок 3. Рисунок 3. Гистопатологические аномалии в образцах сердца от двух субъектов с укороченными мутациями TTN .
Образцы из межжелудочковой перегородки сердца у двух субъектов (панель А и панель В, гематоксилин и эозин) показывают ядра миоцитов с аномальными морфологическими характеристиками (стрелки).
Субъекты с укороченными мутациями TTN имели дилатационную кардиомиопатию, которая обычно не сопровождалась поражением проводящей системы или скелетных мышц (таблицы 9, 10 и 11 в дополнительном приложении), хотя явная скелетная миопатия наблюдалась у одного субъекта с гомозиготным Мутация сплайсинга TTN .Гистопатологические характеристики образцов сердца от субъектов с укороченными мутациями TTN были типичны для идиопатической дилатационной кардиомиопатии. В некоторых срезах были обнаружены очаги миоцитов с причудливой звездчатой ядерной морфологией, которые лучше всего видны на поперечном срезе (рис. 3). Электронно-микроскопическое исследование одного образца, полученного при вскрытии, показало неповрежденные саркомерные структуры (рис. 6 в дополнительном приложении).
Теплофизические свойства некоторых ключевых твердых тел: обновление
Г. К. Уайт и М. Л. Минджес (ред.), Теплофизические свойства некоторых ключевых твердых тел , Бюллетень CODATA № 59 (Пергамон, Оксфорд, 1985).
Google Scholar
Д. Л. Мартин, Rev. Sci. Инструм. 58 :639 (1987).
Артикул ОБЪЯВЛЕНИЯ Google Scholar
К. Н. Марш (ред.), Рекомендованные эталонные материалы для реализации физико-химических свойств , раздел 9 (Blackwell Scientific, Oxford, 1987).
Google Scholar
F. Righini, J. Spisiak, G.C. Bussolino, and A. Rosso, High Temp.-High Press. 25 :193 (1994).
Google Scholar
М. Окаджи, Междунар. Дж. Термофиз. 9 :1101 (1988).
Дж. Термофиз. 9 :1101 (1988).
Артикул Google Scholar
А. П. Мюллер и А. Сезаирлиян, Междунар. Дж. Термофиз. 11 :619 (1990).
Артикул Google Scholar
Р. Б. Робертс, Ф. Ригини и Р. К. Комптон, Фил. Маг. В52 :1147 (1985).
Google Scholar
D.W. Osborne, H.E. Flotow, and F. Schreiner, Rev.науч. Инструм. 38 :159 (1967).
Артикул ОБЪЯВЛЕНИЯ Google Scholar
GT Furukawa, WG Saba, and ML Reilly, NSRDS-NBS 18: Критический анализ литературных данных по теплоемкости и оценка термодинамических свойств меди, серебра и золота от 0 до 300 K (Правительственная типография США, Вашингтон, округ Колумбия, 1968 г. ).
).
Google Scholar
J. C. Holste, T. C. Cetas и C. A. Swenson, Rev. Sci. Инструм. 43 :670 (1972).
Артикул ОБЪЯВЛЕНИЯ Google Scholar
G.K. White and S.J. Collocott, J. Phys. хим. Ссылка Данные 13 :1251 (1984).
ОБЪЯВЛЕНИЕ Статья Google Scholar
р.А. Роби, Б. С. Хемингуэй и У. Х. Уилсон, J. Res. геол. Surv. 4 :631 (1976).
Google Scholar
R. Hultgren, PD Desai, DT Hawkins, M. Gleiser, KK Kelley, and DD Wagman, Selected Values of the Thermodynamic Properties of the Elements (Am. Soc. Metals, Metals Park, OH, 1973).
Google Scholar

А. С. Добросавлевич и К. Д. Маглич, High Temp.-High Press 23 :129 (1991).
Google Scholar
N. E. Phillips, Критический. Преподобный Сол. гос. науч. 2 :467 (1971).
Google Scholar
A. Cezairliyan and J.L. McLure, J. Res. Натл. Бур. Стенд . А78 :1 (1974).
Google Scholar
Ю. С. Тулукян и К. Ю. Хо (ред.). Теплофизические свойства вещества, тома. 1–13 (Пленум Пресс, Нью-Йорк, 1970–1977).
Google Scholar
P. D. Desai, J. Phys. хим. Ссылка Данные 15 :967 (1986).
ОБЪЯВЛЕНИЕ Статья Google Scholar

LW Shacklette, Phys. Версия В9 :3789 (1974).
Артикул ОБЪЯВЛЕНИЯ Google Scholar
Р. К. Кирби, Т. А. Хан и Б. Д. Ротрок, в Справочник Американского института физики, 3-е изд. (Макгроу-Хилл, Нью-Йорк, 1972 г.), гл. 4.
Google Scholar
Дж. Лиз и А. Э. Лорд, Дж.заявл. физ. 39 :3986 (1968).
Артикул ОБЪЯВЛЕНИЯ Google Scholar
М. Л. Рейли и Г. Т. Фурукава, NBS, Вашингтон, округ Колумбия, личное сообщение (1978).
Орлова М.П. и Королев Ю.А., Рос. Дж. Физ. хим. 52 :2756 (1978).
Google Scholar
Д. А. Дитмарс, С. Исихара, С. С. Чанг, Г. Бернштейн и Э. Д. Вест, J. Res. Натл. Бу. Стоять. 87 :159 (1982).
Д. Вест, J. Res. Натл. Бу. Стоять. 87 :159 (1982).
Google Scholar
R. Castanet, High Temp.-High Press. 16 :449 (1984).
Google Scholar
G.K. White and R.B. Roberts, High Temp.-Высокий пресс. 12 :311 (1980).
Google Scholar
F. R. Kroeger and C. A. Swenson, J. Appl. физ. 48 :853 (1977).
Артикул ОБЪЯВЛЕНИЯ Google Scholar
C. A. Swenson, J. Phys. хим. Ссылка Данные 12 :179 (1983).
ОБЪЯВЛЕНИЕ Статья Google Scholar

C. A. Swenson, Университет штата Айова, личное сообщение (1990 г.).
Y. Okada and Y. Tokumaru, J. Appl. физ. 56 :314 (1984).
Артикул ОБЪЯВЛЕНИЯ Google Scholar
К. Г. Лайон, Г. Л. Сэлинджер, К. А. Свенсон и Г. К. Уайт, Дж.заявл. физ. 48 :865 (1977).
Артикул ОБЪЯВЛЕНИЯ Google Scholar
S. J. Bennett, Тепловое расширение 8 , Т. А. Хан, изд., (Plenum Press, New York, 1981), p. 235.
Google Scholar
H. Ibach, Phys. Статус солиди 31 :625 (1969).
Google Scholar
Р. Б. Робертс, J. Phys. Д. 14 :L163 (1981).
Артикул ОБЪЯВЛЕНИЯ Google Scholar
G.K. White and R.B. Roberts, High Temp.-High Press. 15 :321 (1983).
Google Scholar
R.A. Matula, J. Phys. хим. Ссылка Данные 8 :1147 (1979).
ОБЪЯВЛЕНИЕ Статья Google Scholar
J. Bass, Landolt-Börnstein, New Series, Vol. III/15a, Metals: Electronic Transport Phenomena (Springer-Verlag, Berlin, 1982), p. 1.
Google Scholar
T.K. Chu and C.Y. Ho, CINDAS Report 60, а также T.K. Chu, P.D. Desai, and C.Y. Ho, CINDAS Report 71 (Purdue Research Foundation, 1983).
Г. К. Уайт, в Теплопроводность 17 , Дж. Г. Хуст, изд. (Пленум, Нью-Йорк, 1981), с. 95.
95.
Google Scholar
Л. Бесли и Р. К. Кемп, Криогеника 18 :497 (1978).
Артикул Google Scholar
JG Hust, High Temp.-High Press. 8 :377 (1976).
Google Scholar
Дж. Г. Хаст и П. Дж. Джарратано, Специальная публикация NBS 260-52 (Вашингтон, округ Колумбия, 1975).
M. L. Minges, Int. J. Тепломассообмен 17 :1365 (1974).
Артикул Google Scholar
Дж. Н. Манди, Фил. Маг. А46 :345 (1982).
ОБЪЯВЛЕНИЕ Google Scholar
R. Berman, Теплопроводность в твердых телах (Clarendon Press, Oxford, 1976).
Google Scholar
P.G. Klemens, Solid State Phys. 7 :1 (1963).
Артикул Google Scholar
А.Cezairliyan и Y.S. Touloukian, в Достижения в теплофизических свойствах при экстремальных температурах и давлениях , S. Gratch, ed. (ASME, Нью-Йорк, 1965), с. 301.
Google Scholar
Г. Э. Чайлдс, Л. Дж. Эрикс и Р. Л. Пауэлл, Теплопроводность твердых тел при комнатной температуре и ниже , Монография NBS № 131 (Министерство торговли США, Вашингтон, округ Колумбия, 1973).
Google Scholar
Дж. Г. Хаст и А. Б. Ланкфорд, Теплопроводность алюминия, меди, железа и вольфрама от 1 К до точки плавления , Внутренний отчет NBS 84-3007 (Министерство торговли США, Боулдер, 1984).
Google Scholar
C.Y. Ho, R.W. Powell, and P.E. Liley, J. Phys. хим. Ссылка Дата 3 :доп. № 1 (1974).
Дж. Г. Хуст и П.Дж. Джарратано, NBS Spec. Опубл. № 260-50 (Министерство торговли США, Вашингтон, округ Колумбия, 1975 г.).
Google Scholar
C. F. Lucks, J. Test. оценка 1 :Нет. 5 (1973).
Дж. В. Кристиан, Дж. П. Ян, В. Б. Пирсон и И. М. Темплтон, Proc. Рой. соц. А 254 :213 (1958).
ОБЪЯВЛЕНИЕ Google Scholar
G. Borelius, W.H. Keesom, C.H. Johansson, and J.O. Linde, Proc. Рой. акад. Амстердам 35 :10 (1932) и более ранние статьи, цитируемые там.
Google Scholar
J. Nystrom, Ark. Мат. Астр. Фис. 34A :Нет. 27 (1948).
А. А. Рудницкий, Термоэлектрические свойства благородных металлов и их сплавов , Перевод USAEC-tr-3724 (1948).
J. J. Lander, Phys. Ред. 74 :479 (1948).
Артикул ОБЪЯВЛЕНИЯ Google Scholar
Н. Кьюсак и П. Кендалл, Proc. физ. соц. 72 :898 (1958).
Артикул Google Scholar
М. Дж. Лаубиц, Кан. Дж. Физ. 47 :2633 (1969).
ОБЪЯВЛЕНИЕ Google Scholar
R. P. Huebener, Phys. Ред. А140 :1834 (1965).
Артикул ОБЪЯВЛЕНИЯ Google Scholar
J. P. Moore and R. S. Graves, J. Appl. физ. 44 :1174 (1973).
Артикул ОБЪЯВЛЕНИЯ Google Scholar
J.G. Cook, M.J. Laubitz, and M.P. van der Meer, J. Appl. физ. 45 :510 (1974).
Артикул ОБЪЯВЛЕНИЯ Google Scholar
C.A. Rahm and R.D. Barnard, J. Phys. Д 8 :L129 (1975).
Артикул ОБЪЯВЛЕНИЯ Google Scholar
Р. Б. Робертс, Фил. Маг. 36 :91 (1977).
ОБЪЯВЛЕНИЕ Google Scholar
Р. Б. Робертс, Фил. Маг. 43 :1125 (1981).
Google Scholar
Анализ вариации количества копий GK Rat выявил новые предполагаемые локусы предрасположенности к диабету 2 типа
Abstract
Были предприняты большие усилия для поиска генов, ответственных за диабет 2 типа (СД2), но в результате у людей было найдено только около 20 генов из-за его сложности и гетерогенности.Крыса GK, спонтанная модель T2D, предлагает нам превосходную возможность поиска большего количества диабетических генов. Используя технологию сравнительной гибридизации генома (aCGH), мы идентифицировали 137 областей неповторяющихся вариаций числа копий (CNV) у крыс GK при использовании нормальных крыс Wistar в качестве контроля. Эти области CNV (CNVR) охватывают примерно 36 Мб нуклеотидов, что составляет около 1% всего генома. Объединив информацию из аннотаций генов и знаний о заболеваниях, мы всесторонне исследовали CNVR для поиска новых генов T2D.В результате мы отдали предпочтение 16 предполагаемым генам, кодирующим белок, и двум генам микроРНК ( rno-mir-30b и rno-mir-30d ) как хорошим кандидатам. Каталог CNVR между крысами GK и Wistar, выявленный в этой работе, служил хранилищем для добычи генов, которые могут играть роль в патогенезе T2D. Более того, наши усилия по использованию методов биоинформатики для определения приоритетности хороших генов-кандидатов предоставили более конкретный набор предполагаемых кандидатов. Эти результаты будут способствовать изучению генетической основы СД2 и, таким образом, прольют свет на его патогенез.
Каталог CNVR между крысами GK и Wistar, выявленный в этой работе, служил хранилищем для добычи генов, которые могут играть роль в патогенезе T2D. Более того, наши усилия по использованию методов биоинформатики для определения приоритетности хороших генов-кандидатов предоставили более конкретный набор предполагаемых кандидатов. Эти результаты будут способствовать изучению генетической основы СД2 и, таким образом, прольют свет на его патогенез.
Образец цитирования: Ye Z-Q, Niu S, Yu Y, Yu H, Liu B-H, Li R-X, et al. (2010) Анализ вариации количества копий крыс GK выявил новые предполагаемые локусы предрасположенности к диабету 2 типа. ПЛОС ОДИН 5(11): е14077. https://doi.org/10.1371/journal.pone.0014077
Редактор: Stein Aerts, University of Leuven, Бельгия
Получено: 14 июля 2010 г.; Принято: 31 октября 2010 г.; Опубликовано: 23 ноября 2010 г.
Авторские права: © 2010 Ye et al.Это статья с открытым доступом, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Финансирование: Эта работа была поддержана грантами Национальной программы исследований и разработок в области высоких технологий Китая «863» (2009AA022710), Национальной программы фундаментальных исследований «973» (2010CB529206), Национального фонда естественных наук Китая. (30770497, 30800641), Китайский фонд докторантуры (200669), Исследовательская программа Академии наук Китая (KSCX2-YW-R-112), Шанхайский комитет по науке и технологиям (2009B105) и SA-SIBS (Sanofi Aventis — Шанхайский институт биологических наук) Стипендиальная программа.Спонсоры не участвовали в разработке исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявили об отсутствии конкурирующих интересов.
Введение
Диабет 2 типа (СД2), официально известный как инсулиннезависимый диабет, составляет около 90% из 180 миллионов случаев диабета во всем мире [1]. Это хроническое и сложное заболевание, характеризующееся дефектами секреции инсулина бета-клетками островков поджелудочной железы и действием инсулина в периферических тканях, предположительно обусловлено комбинированным действием множества генетических факторов [2], [3].За последние два десятилетия были предприняты большие усилия, включая традиционное картирование генов-кандидатов и недавние высокопроизводительные полногеномные ассоциативные исследования, чтобы раскрыть генетическую основу СД2, и было обнаружено около 20 генов СД2 человека [3]–[8] и число родственных локусов в геномах человека, мыши и крысы [9], [10]. Однако точный молекулярный патогенез этого гетерогенного заболевания остается плохо изученным, и ожидается, что будет обнаружено больше генов, связанных с СД2.
Крыса Goto-Kakizaki (GK), не страдающая ожирением животная модель T2D, была получена путем повторного инбридинга непереносимых к глюкозе крыс Wistar [11].Во время своего развития крысы GK спонтанно страдают от снижения массы бета-клеток и резистентности к инсулину и, таким образом, предоставляют реальную возможность для поиска восприимчивых локусов, изучения патогенеза и разработки терапевтических стратегий [12], [13]. Несколько анализов локуса количественных признаков (QTL) на этой модели уже идентифицировали ряд геномных локусов, несущих восприимчивые варианты [14]–[16].
В то время как большинство исследований генетической изменчивости, связанных с заболеванием, были сосредоточены на отдельных нуклеотидных последовательностях, крупномасштабные изменения, такие как вариации числа копий (CNV), обычно определяемые как различия в числе копий участков ДНК размером более 1 кб, также были связаны с десятками заболевания человека [17].Среди наиболее известных случаев — ассоциация синдрома Дауна с дополнительной копией хромосомы 21, идентифицированной с помощью технологии кариотипирования [18]. Новые высокопроизводительные подходы, такие как сравнительная гибридизация генома на основе массивов (массив CGH или aCGH), позволили идентифицировать CNV во всем геноме [19]–[21] и обнаружили, что CNV широко распространены вдоль хромосом. Обнаружено, что некоторые из CNV связаны со сложными заболеваниями, включая нейропсихиатрические, аутоиммунные заболевания и т. д. [22]–[25], но связь CNV с СД2 остается в значительной степени неиспользованной, за исключением того, что совсем недавнее исследование подтвердило значение ранее идентифицировали человеческий ген TSPAN8 [26].
В этой работе мы провели полногеномный скрининг CNV между GK (модель T2D) и крысой Wistar (дикий тип) с использованием массива CGH. Был идентифицирован неизбыточный набор областей CNV общей длиной около 36 млн. п.н., включая несколько новых локусов предрасположенности к СД2, включающих 16 генов, кодирующих белок ( Il18r1 , Cyp4a3 , Sult2a1 , Sult2a2 , Sult2a2 , Sult2a2 , , NOS2 , PSTPIP1 , UGT2B , UXS1B1 , RT1-A1 , RT1-A1 , RT1-A3 , RT1-DB1 , RT1-N1 , RT1-N1 , RT1-N3 , RT1 -O и RT1-S2 ) и два гена микроРНК ( rno-mir-30b и rno-mir-30d ).На данный момент это первое исследование СД2 у крыс GK с точки зрения изменения количества копий в масштабе всего генома, и предполагается, что CNV, идентифицированные у крыс GK, проливают свет на генетическую основу и патогенез СД2.
Результаты
Обработка данных массива
Сравнение между крысами GK и Wistar имело три биологических повтора («прямой») с заменой красителя («обратный») для каждого, в результате чего было получено 6 массивов. Данные массива обрабатывали в соответствии с процедурами, описанными в разделе «Материалы и методы».На этапе контроля качества набор данных из небольшой области шестого чипа включает 1278 точек (∼0,5%), в дополнение к 152, 157, 147, 143, 155 и 401 рассеянной точке каждого чипа соответственно ( ∼0,08% в среднем), были отфильтрованы из-за плохого качества. Чтобы обнаружить предвзятость маркировки красителем, мы иерархически сгруппировали все 6 чипов на основе отношений логарифмов (значения M). Оказалось, что все «прямые» чипы были четко отделены от «обратных», что противоречило ожиданию того, что пара замены красителя должна быть сгруппирована вместе (рис. S1A).Используя линейную модель, мы обнаружили, что около 23,7% пятен показали значительное смещение красителя ( p <0,05). После исправления смещения красителя новый результат кластеризации совпал с ожидаемым (рис. S1B). Три пары замены красителя привели к трем наборам значений M, каждый из которых соответствовал одному повтору GK. Затем хромосомы были разделены в соответствии со сглаженными значениями M зондов, расположенных на них, процесс, формально называемый сегментацией. Необработанные интенсивности и обработанные данные были депонированы в Gene Expression Omnibus NCBI [27] и доступны через GEO Series номер доступа GSE21387.
Идентификация CNVR
В соответствии с нашим определением областей CNV (CNVR, см. «Материалы и методы»), мы идентифицировали в среднем около 101 CNVR у каждой крысы GK, охватывающих примерно 26 миллионов пар оснований, , т.е. , около 1% генома крысы (таблица S1). и S2), порядок величины которых был таким же, как и общая длина опубликованных CNVR крыс (22 Мб и 15,5 Мб на двух разных платформах) [21]. Сравнение CNVR, идентифицированных из 3 образцов, показало, что большинство из них (более 70%) были одинаковыми среди людей (таблицы S2 и S3).Затем мы объединили CNVR из всех 3 образцов в окончательный неизбыточный набор, состоящий из 137 CNVR, охватывающих 36,31 Мб (таблица S4 и столбец «Неизбыточные» в таблице S2). Последующие исследования были основаны на этом неизбыточном наборе данных.
Мы нанесли CNVR GK/Wistar вдоль каждой хромосомы (рис. 1) и обнаружили, что они были неравномерно распределены с крайними случаями, когда хромосомы 12 и 18 не содержали CNVR, а хромосомы 7 и 15 содержали больше CNVR, чем случайные (4,5 Мб). и 2.7 МБ идентифицировано против только 1,8 МБ и ожидаемых 1,4 МБ соответственно). Неравномерный характер распределения CNVR был в некоторой степени похож на предыдущий отчет о CNVR крыс [21].
Рис. 1. Хромосомное распределение CNVR GK/Wistar.
Зеленые столбцы слева и красные столбцы справа от хромосомных осей представляют «потерю» и «прирост» CNV соответственно. Хромосома «Un» представляет собой псевдохромосому, состоящую из контигов, которые нельзя с уверенностью сопоставить с конкретной хромосомой.
https://doi.org/10.1371/journal.pone.0014077.g001
Расследование CNVR
Чтобы выявить генетические вариации, лежащие в основе фенотипических различий между крысами GK и Wistar (, т. е. , диабетические и недиабетические), мы исследовали CNVR, изучив их перекрытие с различными геномными особенностями, такими как гены, кодирующие белок, и микроРНК. Геномные особенности, охватываемые GK/Wistar CNVR, послужили ценным хранилищем для изучения генетических факторов, которые играют роль в патогенезе T2D посредством измененных чисел копий и, следовательно, аномальных уровней экспрессии.Для гена CNV, о котором уже сообщалось, что он связан с T2D, наша работа может помочь выяснить его основные механизмы, , т.е. , эффект дозы гена через изменение числа копий. Что еще более важно, гены CNV, о которых не было известно, что они связаны с СД2, могут содержать новых кандидатов, и предполагалось, что будет многообещающе отделить их от неродственных путем объединения предшествующих биологических знаний с использованием методов биоинформатики.
Генное и межгенное содержание CNVR
Генные и межгенные области в CNVR GK/Wistar определяли в соответствии с хромосомным расположением крысиных генов, полученным из NCBI Entrez Gene.В совокупности области размером 3,22 Мб были аннотированы Entrez Gene, что составляет всего 8,87% всех CNVR, что намного меньше, чем доля областей генов во всем геноме (21,35%, таблица 1). Казалось, что вариации числа копий будут преимущественно находиться в межгенных областях. Далее мы провели случайное моделирование для проверки статистической значимости (см. «Материалы и методы») и обнаружили, что величина перекрытия между рандомизированными интервалами и генами была значительно больше, чем между реальными CNVR и генами (7.84 Мб ожидаемо случайным образом со стандартным отклонением 1,13 Мб по сравнению с 3,22 Мб фактически, p <1e-10), что подтверждает наш вывод о предпочтении CNVR межгенным областям. Частично это можно объяснить очищающим отбором, который, вероятно, действовал на CNV GK/Wistar в процессе селекционного разведения. Вариации в большинстве областей генов могут с большей вероятностью оказывать вредное воздействие, чем в межгенных областях, и, таким образом, более сильное негативное давление отбора на области генов может привести к более низкой наблюдаемой частоте CNV в областях генов, чем в межгенных областях.
Функциональный анализ генов CNV
Всего было идентифицировано 62 и 72 гена Entrez в CNVR с «приобретением» и «потерей» соответственно (таблицы S4 и S5). Мы проверили, содержат ли эти гены какие-либо из тех, о которых ранее сообщалось, что они связаны с СД2. Был собран список из 425 известных генов, связанных с СД2, из опубликованных источников (см. Материалы и методы) и проведено сравнение с этими 134 генами GK/Wistar CNV, но ничего общего не было обнаружено. Этот результат показал, что потенциальными факторами CNV, лежащими в основе диабетического патогенеза крыс GK, могут быть другие гены, связь которых с T2D не наблюдалась.Это могут быть гены, которые были аннотированы в NCBI Entrez Gene, или даже локусы, которые ранее никогда не идентифицировались. В этой работе мы сосредоточились на добыче кандидатов из известных генов (то есть 134 генов CNV) в соответствии с вышеупомянутой стратегией принятия предварительных знаний.
Сначала мы использовали базу знаний путей KEGG, чтобы идентифицировать потенциальных новых кандидатов, которые, как предполагается, связаны с характеристиками диабета. После сопоставления 134 генов CNV с KEGG мы обнаружили, что с этими генами связан 41 путь (таблица S6).Затем для дальнейшего анализа были отобраны гены CNV, попадающие в пути, связанные с T2D (таблица 2). Поскольку T1D имеет некоторые общие характеристики с T2D, и ранее сообщалось об участии метаболизма серы в диабете [28], [29], гены CNV ( RT1-A1 , RT1-A3 , RT1-Db1 , RT1-N1 , RT1-N1 , RT1-N3 , RT1-O , RT1-S2 , Sult2Al1 , Sult2A1 , Sult2A1 и Sult2A2 ) в путях «Тип I Diabetes Mellitus (04940)» и приоритет отдавался «обмену серы (00920)».Хорошо известно, что метаболизм сахаров или жирных кислот и сигнальные пути PPAR связаны с СД2 [2], и, таким образом, гены CNV ( Uxs1 , Ugt2b и Cyp4a3 ) в путях «метаболизма крахмала и сахарозы» (00500 )», «взаимопревращения пентозы и глюкуроната (00040)», «метаболизм жирных кислот (00071)» и «передача сигналов PPAR (03320)». Мы также обнаружили, что Ugt2b , Cyp4a3 и CNV, связанные с метаболизмом серы ( Sult2a1 , Sult2a2 и Sult2al1 ), перекрываются с T2D QTL Niddm37, эти гены могут обусловливать предрасположенность к T2D.
Мы заметили, что гены GK/Wistar CNV были значительно перепредставлены в путях обонятельной трансдукции (04740), иммунного ответа (05332, 05330, 05320 и 04612) и молекулы клеточной адгезии (04514) (таблица S6). Они напоминали результаты функционального обогащения, о которых ранее сообщалось в генах CNV человека, мыши и крысы, указывая на то, что смещение в сторону этих функциональных категорий может быть общей общей характеристикой генов CNV [19], [21], [30].
Поскольку существуют сложные взаимосвязи между заболеваниями, составляющие «заболевание» [31], заболевания, непосредственно связанные с СД2 («близкие к СД2»), могут помочь понять родство генов GK/Wistar CNV с СД2.Всего было извлечено 1097 генов, близких к СД2, в соответствии с процедурой, описанной в разделе «Материалы и методы». Из-за сложности и гетерогенности диабета мы предположили, что часть этих генов также может в некоторой степени способствовать развитию СД2. Фактически, в списке генов GK/Wistar CNV встречаются пять генов, близких к T2D, включая Pstpip1 , Il18r1 , Sult2a1 , Nos2 и RT1-Db1 (T) Мы также проверили взаимосвязь между ними и крысиными QTL и обнаружили, что они перекрываются с QTL «кровяного давления», «триглицеридов сыворотки», «холестерола сыворотки» или «массы тела».В частности, Pstpip1 , Il18r1 , Sult2a1 перекрывают QTL «инсулинозависимого сахарного диабета (Niddm)».
Существует несколько веб-инструментов для приоритизации генов-кандидатов болезней человека из заданных геномных интервалов [32]. Сравнение между нашими 134 крысиными генами CNV и 103 человеческими генами T2D, приоритетными для Tiffin et al. [33] обнаружил один общий ген, Uxs1 , который также был отсортирован с помощью нашего картирования пути, связанного с T2D.
В совокупности вышеприведенный анализ выявил 16 генов GK/Wistar CNV, предоставив ценную коллекцию наиболее вероятных кандидатов на заболевание, которые должны быть приоритетными для дальнейших экспериментов (таблица S8). Эти гены неравномерно распределены по нескольким разным хромосомам и разным регионам CNV. Среди них все 7 генов RT1 кластеризованы в двух областях хромосомы 20, т.е.Усиление.54 и cnv.gain.53 составляет всего ∼345 Кб в соответствии с таблицей S4), в то время как RT1-N1 , RT1-N3 , RT1-O и RT1-S2 в другом. Однако 3 гена в первом кластере поддерживаются только одним образцом, что снижает нашу уверенность в том, что эти гены могут способствовать предрасположенности к СД2. Три гена Sult2a сгруппированы в области усиления на хромосоме 1 и поддерживаются всеми тремя образцами. Остальные шесть генов распределены по шести различным хромосомам соответственно, причем Uxs1 , Cyp4a3 и Nos2 идентифицированы во всех трех образцах.Эти данные дадут дополнительные подсказки для последующей расстановки приоритетов этих 16 генов.
Ультраконсервативные элементы и микроРНК
Согласно вышеупомянутому анализу, более 90% CNVR были межгенными областями. Хотя межгенные области, а также некодирующие области генов, такие как интроны и UTR, могут не включать столько функциональных признаков, как кодирующие области, недавние исследования показали, что некоторые из некодирующих областей могут играть важную регуляторную роль.Поэтому мы расширили наше внимание за пределы кодирующих областей, чтобы включить ультраконсервативные элементы и микроРНК.
Ультраконсервативные элементы определяются как участки ДНК (длиной более 200 п.н.), которые чрезвычайно консервативны в ортологичных областях геномов человека, крысы и мыши. Было высказано предположение, что они находятся под сильным очищающим отбором и могут играть важную роль в связывании ДНК, процессинге РНК и регуляции транскрипции [34]. Мы сравнили CNVR GK/Wistar и 481 опубликованный ультраконсервативный элемент (126.всего 7 т.п.н., сопоставленных со сборкой генома крысы rn4), но перекрытия обнаружено не было, в то время как ∼1,5 т.п.н. можно было ожидать случайно ( p = 0,027), что согласуется с выводами, сделанными в других исследованиях [21], [35] . Эти результаты в сочетании с вышеуказанным генным и межгенным содержанием CNVR подтверждают мнение о том, что в качестве глобальной тенденции большинство CNV ГК/Вистар были обнаружены в нефункциональных, а не функциональных областях, вероятно, из-за очищающего отбора во время селекционного разведения. .
Принимая во внимание значительный вклад микроРНК в процессы развития и патогенез заболеваний на посттранскрипционном уровне [36]–[37], мы проверили наличие генов микроРНК в наших GK/Wistar CNVR.Сравнивая геномные положения известных генов крысиных микроРНК с геномами GK/Wistar CNVRs, мы обнаружили, что rno-mir-30b и rno-mir-30d были одновременно покрыты областью «прироста» на хромосоме 7 во всех три образца (таблица S9) в области всего 3,8 Кб. Интересно, что несколько QTL T2D и различные соответствующие QTL, включая QTL массы тела, артериального давления и уровня триглицеридов в сыворотке, также были расположены в этой области, что означает, что изменение количества копий этих генов микроРНК может быть коррелировано с этими количественными признаками (рис. 2).В недавней публикации сообщалось, что измененная экспрессия mir-30d в ответ на глюкозу влияет на экспрессию гена инсулина в мышиной Min6, линии островковых клеток поджелудочной железы [38]. Хотя дальнейшие исследования все еще необходимы, мы получили дополнительные доказательства, подтверждающие участие mir-30b/30d в патогенезе T2D посредством изменения количества копий.
Рис. 2. МикроРНК rno-mir-30b и rno-mir-30d , расположенные в QTL T2D.
QTL Niddm (инсулиннезависимый сахарный диабет) 14 и 19 охватывают эти 2 микроРНК.Кроме того, в этой области есть много других QTL, таких как «триглицерид сыворотки 14», «холестерин сыворотки 3», «артериальное давление 181/215/216/265/266», «масса тела 9/12/17», и эти признаки, как известно, связаны с диабетом. Этот рисунок был подготовлен с использованием браузера генома UCSC.
https://doi.org/10.1371/journal.pone.0014077.g002
Для дальнейшего выяснения предполагаемой роли mir-30b/30d мы рассмотрели их предполагаемые цели с помощью MicroCosm [39]. В совокупности для рно-мир-30б и рно-мир-30д было 1868 и 1776 мишеней соответственно.Как и при анализе генов CNV, мы сравнили эти гены-мишени с 425 генами, связанными с T2D. Оказалось, что 39 и 35 мишеней mir-30b и mir-30d встречались в этом списке генов T2D соответственно, и обе были значительно перепредставлены ( p = 0,000273 и 0,00152, подробные мишени перечислены в таблице 3). подтверждая гипотезу об участии mir-30b и mir-30d в T2D. Среди них PPARG и AKT2 и AKT2 (цели MIR-30B ), HNF1B , HNF4A и LMNA (цели MIR-30D ), являются известными генами, привлеченными в T2D или резистентность к инсулину.Затем мы сопоставили эти мишени микроРНК с путями KEGG и обнаружили, что 5(2), 12(6), 10(4), 14(12), 4(5) и 1(3) мишени mir-30b ( mir-30d ) относились к путям «диабета II типа (04930)», «диабета I типа (04940)», «рака поджелудочной железы (05212)», «сигнализации инсулина (04910)», «сигнализации PPAR (03320)». )» и «диабет зрелого возраста у молодых (04950)» соответственно (табл. 4). Более того, несколько путей метаболизма жирных кислот или сахаров (00010, 00030, 00512, 00051, 00071 и 01030) были обогащены этими предсказанными мишенями со значительно низкими значениями p (таблица S10 и таблица S11).Эти результаты предоставили дополнительные доказательства роли mir-30b/30d в патогенезе диабета.
Обсуждение
В этом исследовании мы определили каталог CNVR между крысами GK и Wistar, используя мозаичный массив CGH. Принимая во внимание гипотезу о том, что фенотипические различия между GK и Wistar (диабетическими и недиабетическими) должны быть в значительной степени связаны с их геномными вариациями, мы провели серию биоинформатических функциональных анализов этих GK/Wistar CNVR, чтобы сузить диапазон. возможности для дальнейшего изучения генов-кандидатов СД2.В общей сложности 16 генов, кодирующих белок, и 2 гена микроРНК были выбраны в качестве приоритетных для дальнейшего анализа (таблица S8 и таблица 3), которые могут в комбинации или по отдельности способствовать патогенезу диабета на основе различного количества копий на геномном уровне.
Мы считаем, что список CNVR GK/Wistar является ценным хранилищем для изучения генетических факторов, которые играют роль в патогенезе T2D посредством измененных номеров копий. В биоинформатическом анализе мы сосредоточились на кодирующих белок генах Entrez и микроРНК с известными геномными местоположениями и обнаружили 134 кодирующих белок гена и 2 микроРНК, вовлеченных в GK / Wistar CNVR.Объединив имеющиеся знания о СД2, мы отдали приоритет 16 генам, кодирующим белок, и 2 микроРНК как хорошим кандидатам для дальнейших экспериментов по проверке их вклада в патогенез СД2 с помощью эффекта дозы. Кроме того, CNVR без аннотаций генов Entrez или микроРНК также могут заслуживать дальнейшего изучения. Мы проверили другие аннотации генов из UCSC «KnownGene», «RefGene», «мРНК», «EST» и «EnsemblGene» и обнаружили различные кодирующие сигналы, выходящие за рамки аннотаций генов Entrez (таблица S12).Это могут быть новые гены, но необходимы дополнительные доказательства.
Среди предпочтительно выбранных генов, кодирующих белок, Il18r1 ранее был идентифицирован как ген-кандидат T2D в когорте афроамериканских семей [40]. Ген Cyp4a3 участвует в метаболизме жирных кислот и передаче сигналов Pparg, нарушения которых тесно связаны с патогенезом СД2 [2], [41]. Как упоминалось в результатах, некоторые серосодержащие соединения используются в терапии диабета, хотя терапевтический механизм еще не ясен.Таким образом, идентификация sult2a1 , sult2a2 и sult2al1 перекликается с полезными эффектами серосодержащих соединений, и дальнейшее изучение может пролить свет на механизм.
Мы проанализировали общедоступный набор данных GSE13271 [42], в настоящее время единственный набор данных дифференциальной экспрессии GK/Wistar в NCBI GEO, и идентифицировали 30 наборов дифференциально экспрессируемых генов (DEG) между крысами GK и Wistar, соответствующими различным тканям, временным точкам и кормлению. условия.При сравнении этих DEG с генами GK/Wistar CNV, идентифицированными в этой работе, мы обнаружили, что некоторые из них проявляют согласованность в определенных тканях (таблица S13), в том числе RT1-N1 в мышцах, Ugt2b в печени, RT1- A3 во всех трех тканях (печень, жир и мышцы). Была также несоответствие между CNV и дифференциальным выражением, в том числе Sult2AL1 , Sult2A1 , Sult2A2 , Sult2A2 , UXS1 , RT1-DB1 , CYP4A3 , PSTPIP1 и IL18R1 в определенных условиях.Вообще говоря, разумно предположить, что устойчивые гены могут быть более вероятно вовлечены в T2D, но мы не можем исключать возможность вовлечения других генов.
Было много сообщений о влиянии микроРНК на диабет, но почти все они были сосредоточены на профилях экспрессии, которые в основном связаны с промежуточным процессом развития заболевания [38], [43]–[46]. Здесь мы впервые нашли доказательства того, что микроРНК могут быть связаны с T2D посредством вариации числа копий.Мы предположили, что измененное количество копий mir-30b и mir-30d у крыс GK может способствовать патогенезу T2D. Это может происходить на стадии инициации заболевания: по сравнению с нормальными крысами Вистар, различное число копий mir-30b и mir-30d в GK может приводить к изменению уровня экспрессии на некоторых специфических стадиях развития и в некоторых специфических тканях. а измененная экспрессия mir-30b и mir-30d может затем привести к дисфункции некоторых специфических мишеней, способствуя развитию СД2.Все эти предрасполагающие факторы могут действовать в сочетании, поскольку они вовлечены в это сложное заболевание. В дополнение к вышеупомянутому исследованию экспрессии mir-30d , было несколько других сообщений о профилировании экспрессии, предполагающих участие семейства mir-30 в диабете или адипогенезе [47]-[49]. Мы повторно проанализировали общедоступный набор данных по экспрессии микроРНК GSE13920, который в настоящее время является единственным профилированием микроРНК у крыс GK и Wistar [43], и обнаружили, что уровни экспрессии mir-30b/30d в мышечных клетках резко различались между нормальными крысами и крысами. Крыса T2D (рисунок S2).Однако изменение экспрессии противоречило обнаруженному здесь направлению изменения числа копий. Мы заметили, что существует ген, кодирующий белок, названный Zfat , который расположен в том же коэффициенте усиления CNVR, что и mir-30b и mir-30d . Изучив набор данных GSE13271, мы обнаружили, что Zfat Активация повышалась в печени, но подавлялась в жировых тканях и мышцах (таблица S13). Понижающая регуляция Zfat в мышцах согласуется с таковой у mir-30b и mir-30d , то есть все они не согласуются с усилением CNV, что предполагает необходимость дальнейших исследований для подтверждения этих результатов. и раскрыть подробные механизмы.
Хотя мы стремились найти специфические для диабета варианты, мы заметили, что CNVR GK/Wistar, идентифицированные в этой работе, имеют довольно много общих характеристик с ранее опубликованными исследованиями CNV [19], [21], [30]. Например, наши CNVR из одного образца ГК составляют около 1% всего генома, неслучайно распределены по хромосомам и обогащены генами, связанными с обонятельной трансдукцией и иммунным ответом (таблица S6), что предполагает возможные общие факторы, участвующие в генезисе. или техническое обслуживание CNV.Предпочтение CNVR GK/Wistar межгенным областям соответствовало нескольким опубликованным сообщениям [19], [50], в то время как некоторые другие заявляли об обратном [21], [51]. Это может быть связано с различением различных предметов исследования или другими причинами, не выявленными в настоящее время.
В этом исследовании мы попытались изучить причины T2D у крыс GK на основе данных CNV. По общему признанию, было еще несколько аспектов, не затронутых здесь. T2D является гетерогенным заболеванием и может быть вызвано множеством факторов, включая генетические вариации (точечные вариации, структурные вариации, такие как инверсия, транслокация, небольшие вставки и CNV) и воздействие окружающей среды.Также сообщается, что эпигенетические факторы могут быть связаны с СД2 у крыс GK [12]. В настоящей работе рассматриваются только CNV, и, таким образом, весьма ожидаемы будущие исследования, касающиеся всех этих точек и проверки кандидатов. Дизайн зонда с более высокой плотностью, гибридизация с большим количеством особей GK и аннотация генома крысы с большей точностью улучшат качество данных CNVR и последующего анализа. Будет еще более многообещающим, если технология секвенирования следующего поколения будет принята для обнаружения CNV, поскольку она может не только более точно определять границы CNV, но также обнаруживать инверсии или транслокации, которые иначе не могут быть обнаружены с помощью массива CGH.Кроме того, секвенирование может идентифицировать новые участки последовательности, которые не представлены в текущей сборке эталонного генома, прокладывая более широкий путь к идентификации генов T2D и других заболеваний.
Материалы и методы
Подготовка проб, гибридизация массивов и извлечение данных
Три самца крыс Goto-Kakizaki и 8 самцов крыс Wistar были получены от SLAC Co., Ltd (Шанхай, Китай). Крыс анестезировали формалином в возрасте 8 недель, брали кровь из перикарда и антикоагулировали.Затем выделяли геномную ДНК с использованием набора DNeasy Blood & Tissue Kit (Qiagen, p/n 69504). Все эксперименты на животных были одобрены Комитетом по этике биомедицинских исследований Шанхайского института биологических наук Китайской академии наук (IRB00005813). ДНК от 3 особей крысы GK использовали в качестве тестового образца отдельно, тогда как ДНК, объединенная от 8 крыс Wistar, служила в качестве общего эталона.
Мы использовали геном крысы CGH 244A (Agilent, p/n G4435A) в качестве нашей олиго-платформы aCGH, которая включает около 240 тыс. 60-мерных зондов, расположенных поверх генома крысы со средним расстоянием между зондами, равным 7.9 т.п.н. (4,7 т.п.н. в областях генов). Мечение геномной ДНК и гибридизацию с мозаичными матрицами проводили в соответствии со стандартными протоколами Agilent. Вкратце, 2 мкг геномной ДНК из тестового образца (GK) и 2 мкг из эталона (Wistar) расщепляли с помощью AluI/RsaI и метили случайным праймером, включающим флуоресцентные красители Cy5 (красный) и Cy3 (зеленый). Исследуемый и эталонный образцы были совместно гибридизованы с микрочипом Agilent 244A в камере для гибридизации (Agilent, номер по каталогу G2545A) при 65 градусах Цельсия и 20 об/мин в течение 40 часов.Для каждого образца проводили мечение с заменой красителя и гибридизацию. При «прямой» гибридизации испытуемый и эталонный образцы были окрашены Cy5 и Cy3 соответственно, тогда как при соответствующей «обратной» гибридизации испытуемый и эталонный образцы были окрашены Cy3 и Cy5 соответственно.
После гибридизации и промывки массивы визуализировали с помощью сканера высокого разрешения US80803205 (Agilent, номер по каталогу G2505B). Интенсивность флуоресценции была извлечена с использованием программного обеспечения Agilent Feature Extraction и использована для последующих процедур обработки.
Обработка данных
Данные обрабатывались в среде программирования R (2.9.0) и пакетах snapCGH (1.12.0), limma (1.18.0), MANOR (1.16.0) и DNAcopy (1.18.0) из проекта bioconductor ( http://www.bioconductor.org/), и несколько внутренних скриптов были приняты для контроля качества, нормализации и сегментации. Структура обработки данных была в основном основана на snapCGH [52], и были внесены некоторые необходимые модификации, чтобы соответствовать требованиям контроля качества и схемы замены красителя.Весь конвейер, включающий обработку данных и идентификацию CNVR, показан на рисунке 3. Интенсивности сигналов от двух каналов (Cy5 и Cy3) были преобразованы в форму отношений логарифмов (значения M), а затем были введены в MANOR для оценки качества. особенно для настройки потенциального глобального градиента и обнаружения областей чипа с низким качеством [53]. После этого все значения М были нормализованы с помощью «лёссовых» коэффициентов, рассчитанных по значениям М между -1 и 1 («взвешенный лёсс») [54], а значения из «обратных» гибридизаций были умножены на -1, чтобы сделать их сопоставимыми. с теми из «передовых» ( i.е. , значения M для Wistar и GK, преобразованные в значения M для GK и Wistar).
Далее мы разработали линейную модель, используя limma для коррекции смещения красителя, специфичного для пятна [55]. Для каждой характерной точки у нас было 6 M значений, соответствующих 6 массивам. В нашей модели каждое измеренное значение M было записано как: где i принимали значения от 1 до 6, представляя разные массивы. Элемент α использовался для моделирования вклада геномных различий между крысами GK и Wistar, в то время как элемент β предназначался для систематической ошибки красителя.Пункт и представлял собой вклад других факторов, включая разницу между особями GK и случайные ошибки. Во всех гибридизациях ( i = 1, …, 6) GkWis принимал значение 1. Краситель принимал значение 1 для «прямых» гибридизаций (i = 1, 2, 3) и −1. для «обратных» ( и = 4, 5, 6), что свидетельствует о разной направленности смещения красителей при «прямой» и «обратной» гибридизациях. Коррекция смещения красителя заключалась в вычитании компонента β·Dye i из значений M.Минимизируя сумму квадратов e , мы смогли получить оценки α и β в лимме. Дендрограммы, использованные для иллюстрации необходимости коррекции смещения красителя для конкретных точек, были созданы путем иерархической кластеризации с использованием метода минимальной дисперсии Уорда с использованием евклидовых расстояний.
После коррекции смещения красителя 6 массивов были объединены в 3 независимых набора данных, соответствующих 3 образцам ГК. Значения M из повторов зонда также усредняли.Затем была проведена сегментация с использованием пакета DNAcopy, целью которого является фрагментация хромосом на интервалы в соответствии со сглаженными значениями M зондов, расположенных рядом с геномом, , т.е. , статусом количества копий соответствующих областей генома [56].
Идентификация CNVR
В этой работе область CNV (CNVR) была определена на основе сглаженных значений M: трех или более последовательных зондов, все значения M которых были больше 0,5 («усиление») или все меньше -0.5 («потеря») очерчена основная область, которая расширяется дополнительно на 5 Кб с обеих сторон, чтобы определить CNVR. При сравнении двух диплоидов значение M, равное 0,5, предполагает, что один из двух аллелей дублируется (log 2 (3/2)≈0,58). Вот почему 0,5 было установлено как пороговое значение М. Расширение CNVR на 5 Кб с обеих сторон было связано с тем, что 5 Кб соответствуют половине медианного расстояния между последовательными зондами на платформе Agilent 244A, примерно представляя покрытие зонда с одной стороны.
При объединении CNVR нескольких образцов в неизбыточный набор критерием было то, что область, которая была обнаружена хотя бы 1 образцом, должна была остаться в конечном наборе, а перекрывающиеся области должны быть объединены, аналогично «объединению» в набор операций. Хотя «объединение» 3 образцов неизбежно приводило к более высокому уровню ложноположительных результатов идентификации CNVR, мы по-прежнему предпочитали высокий охват высокой точности. Геномные координаты CNVR относили к сборке UCSC rn4 (на основе RGSC 3.4), а их хромосомное распределение строили с помощью Caryoscope [57].
Расследование CNVR
Анализы также проводились в среде программирования R. Хромосомные местоположения генов NCBI Entrez были получены из пакета org.Rn.eg.db (2.2.11) в Bioconductor. Все координаты геномных особенностей, принятые в этой работе, были отнесены к сборке UCSC rn4 в соответствии с координатами CNVR. Гены CNV идентифицировали по перекрытию геномных интервалов, i.е. , если какой-либо из CNVR GK/Wistar перекрывался, он учитывался. При расчете длины участков генов в CNVR или во всем геноме перекрывающиеся участки генов объединялись в неповторяющиеся интервалы, чтобы избежать дублирования подсчета. .
Случайное моделирование для оценки статистической значимости предпочтения CNVR межгенным областям было разработано следующим образом. То же количество геномных интервалов, что и у CNVR (137 в этом исследовании), было случайным образом выбрано из хромосом крыс, и их длины также были такими же, как у соответствующих CNVR.Суммарная длина перекрытий между ними и областями гена Entrez затем рассчитывалась как статистика. После 1000 циклов этого процесса было получено эмпирическое распределение, представляющее величины перекрытия между областями гена и случайными геномными интервалами. И, таким образом, можно было оценить вероятность того, что длина перекрытия меньше, чем между реальными CNVR и областями гена (3,22 Мб в этом исследовании).
Список генов, связанных с СД2, был составлен на основе статьи, в которой собрано 172 гена, связанного с СД2 человека [10], и базы данных СД2-БД [9], которая содержала 330, 60 и 36 генов человека, мыши и крысы соответственно.Используя NCBI HomoloGene Release 64 (http://www.ncbi.nlm.nih.gov/homologene), мы получили соответствующие крысиные гомологи генов человека и мыши. В общей сложности оказалось, что 425 неперекрывающихся крысиных генов составляют список генов, связанных с СД2.
В дополнение к org.Rn.eg.db (2.2.11) пакеты биопроводников, включая KEGG.db (2.2.11) и GOstats (2.10.0), использовались для получения сопоставления генов с путями и для тестирования значение обогащения путей. Набор генов, близких к T2D, был получен из исследования «болезни» человека [31], где все заболевания были организованы во взаимосвязанную сеть.Мы собрали все заболевания, которые напрямую связаны с СД2 в этой сети, а затем все гены, соответствующие этим заболеваниям, были извлечены как набор генов, близких к СД2. Крысиные гомологи человеческих генов, близких к T2D, также были получены из выпуска 64 NCBI HomoloGene, в результате чего было получено 1097 крысиных генов, близких к T2D. Данные крысиного QTL были получены из источника RGD [58], и их перекрытие с генами CNV было проверено путем ручной навигации в браузере генома UCSC [59]. Набор генов Tiffin T2D человека [33] первоначально был представлен идентификаторами генов Ensembl и был преобразован в идентификаторы генов Entrez с использованием org.Hs.eg.db (2.2.11). Человеческие гомологи крысиных генов CNV также были получены с использованием HomoloGene выпуска 64, что сделало возможным сравнение между CNV генами и набором данных Tiffin.
Расположение ультраконсервативных элементов в геноме крысы (сборка rn4) определяли с помощью BLAT [60]. 481 опубликованный элемент привел к 484 позициям из-за неуникального сопоставления. Моделирование для оценки значимости перекрытия между CNVR и ультраконсервативными элементами проводилось в соответствии с предыдущей процедурой для перекрытия между CNVR и областями гена.
крысиных микроРНК вместе с их геномными координатами на основе сборки rn4 были загружены с сайта miRBase (http://www.mirbase.org/, выпуск 14) [61]. Предсказанные мишени зрелых микроРНК были получены от MicroCosm (http://www.ebi.ac.uk/enright-srv/microcosm/htdocs/targets/v5/) на основе алгоритма miRanda [39]. Поскольку rno-mir-30b был обработан в две зрелые формы, rno-miR-30b-3p и rno-miR-30b-5p , их мишени были объединены для дальнейшего анализа; и то же самое было с rno-mir-30d , где мишени rno-miR-30d и rno-miR-30d* были объединены.Цели в MicroCosm, представленные в виде идентификаторов транскриптов Ensembl, были преобразованы в идентификаторы генов Entrez с использованием карт, предоставленных пакетом org.Rn.eg.db. Значимость обогащения мишеней 425 генами, связанными с СД2, рассчитывали в гипергеометрическом распределении, принимая все гены в пакете «org.Rn.eg.db» в качестве фона. Картирование путей и анализ обогащения мишеней проводили так же, как и для генов CNV, с использованием пакета GOstats.
Суперсерия GSE13271 содержит три набора данных экспрессии: GSE13268 (жировая ткань), GSE13269 (мышцы) и GSE13270 (печень), все из которых профилировали экспрессию генов у крыс GK и Wistar при нормальной диете и диете с высоким содержанием жиров и в 5 временных точках. , что приводит к 30 условиям.Для генов с более чем одним набором зондов мы сохранили тот, который чаще всего ассоциировался с самым высоким уровнем экспрессии. Если оставалось еще более одного набора зондов, интенсивности оставшихся наборов зондов усреднялись, чтобы представить уровень экспрессии этого гена. Затем мы выполнили тест t для идентификации дифференциально экспрессируемых генов между образцами GK и Wistar и рассчитали логарифм по основанию 2 кратных изменений для уровней экспрессии. Коррекция FDR с использованием метода BH [62] была принята для корректировки необработанных p -значений нескольких проверок гипотез и 0.2 был выбран в качестве порога, что означает, что более 80% идентифицированных генов действительно дифференциально экспрессируются. Что касается набора данных экспрессии микроРНК GSE13920, мы просто посмотрели на среднюю интенсивность сигнала после удаления среднего фонового шума для каждого зонда mir-30b и mir-30d .
Дополнительная информация
Рисунок S1.
Эффект коррекции смещения красителя. Образцы были сгруппированы в соответствии с соответствующими значениями M до (A) и после (B) исправления систематических ошибок, вызванных смещением красителя, соответственно.GK1′, GK2′ и GK4′ представляют собой соответствующие «обратные» гибридизации для GK1, GK2 и GK4 соответственно.
https://doi.org/10.1371/journal.pone.0014077.s001
(0,08 МБ TIF)
Рисунок S2.
Уровни экспрессии mir-30b/30d в мышцах крыс GK и Wistar. Данные были загружены из GEO (GSE13920), и два образца GK и 2 образца Wistar были гибридизованы на 4 одноканальных микроматрицах соответственно. Уровень экспрессии был представлен средней интенсивностью сигнала переднего плана после вычитания средней интенсивности фонового сигнала.Каждый зонд дублируется 3 раза.
https://doi.org/10.1371/journal.pone.0014077.s002
(0,22 МБ TIF)
Благодарности
Мы благодарим доктора Tingqing Guo из Национального института диабета, болезней органов пищеварения и почек США (NIDDK) за полезные обсуждения. Мы также благодарим доктора Штейна Аэртса и двух анонимных рецензентов за их комментарии и предложения.
Вклад авторов
Задумал и спроектировал эксперименты: HSX RZ YXL JRW YYL.Проведены эксперименты: YY RXL. Проанализированы данные: ZQY SN HY BHL YYL. Написал статью: ZQY YYL.
Каталожные номера
- 1. ВОЗ (2008 г.) Информационный бюллетень ВОЗ по диабету № 312.
- 2. Stumvoll M, Goldstein BJ, van Haeften TW (2005)Диабет 2 типа: принципы патогенеза и терапии. Ланцет 365: 1333–1346.
- 3. Прокопенко И., Маккарти М.И., Линдгрен С.М. (2008)Диабет 2 типа: новые гены, новое понимание. Тенденции Жене 24: 613–621.
- 4. Frayling TM (2007) Полногеномные ассоциативные исследования дают новое представление об этиологии диабета 2 типа. Нат Рев Генет 8: 657–662.
- 5. McCarthy MI, Zeggini E (2009)Общегеномные ассоциативные исследования диабета 2 типа. Карр Диаб, представитель 9: 164–171.
- 6. Zeggini E, Weedon MN, Lindgren CM, Frayling TM, Elliott KS, et al. (2007) Репликация сигналов полногеномной ассоциации в образцах Великобритании выявляет локусы риска для диабета 2 типа.Наука 316: 1336–1341.
- 7. Саксена Р., Войт Б.Ф., Лысенко В., Бертт Н.П., де Баккер П.И. и соавт. (2007) Анализ ассоциации всего генома идентифицирует локусы для диабета 2 типа и уровней триглицеридов. Наука 316: 1331–1336.
- 8. Скотт Л.Дж., Молке К.Л., Бонникасл Л.Л., Виллер С.Дж., Ли И. и др. (2007) Полногеномное исследование ассоциации диабета 2 типа у финнов выявило множественные варианты восприимчивости. Наука 316: 1341–1345.
- 9. Агравал С., Димитрова Н., Натан П., Удаякумар К., Лакшми С.С. и др.(2008) T2D-Db: интегрированная платформа для изучения молекулярной основы диабета 2 типа. BMC Genomics 9: 320.
- 10. Хэнкок А.М., Витонски Д.Б., Гордон А.С., Эшель Г., Притчард Дж.К. и соавт. (2008) Адаптация к климату в генах-кандидатах для общих нарушений обмена веществ. PLoS Genet 4: e32.
- 11. Гото Ю., Какидзаки М., Масаки Н. (1975)Спонтанный диабет, вызванный селективным размножением нормальных крыс Вистар. Proc Jpn Acad 51: 80–85.
- 12. Порта Б., Лакраз Г., Кергоат М., Хомо-Деларш Ф., Жируа М.Х. и др.(2009) Бета-клетка GK крысы: прототип больной бета-клетки человека при диабете 2 типа? Mol Cell Endocrinol 297: 73–85.
- 13. Movassat J, Calderari S, Fernandez E, Martin MA, Escriva F, et al. (2007) Диабет 2 типа — вопрос недостаточного неогенеза бета-клеток? Подсказки из крысиной модели GK. Diabetes Obes Metab 9: Suppl 2187–195.
- 14. Gauguier D, Froguel P, Parent V, Bernard C, Bihoreau MT, et al. (1996) Хромосомное картирование генетических локусов, связанных с инсулиннезависимым диабетом у крыс GK.Нат Жене 12: 38–43.
- 15. Галли Дж., Ли Л.С., Глейзер А., Остенсон К.Г., Цзяо Х. и др. (1996)Генетический анализ инсулиннезависимого сахарного диабета у крыс GK. Нат Жене 12: 31–37.
- 16. Granhall C, Park HB, Fakhrai-Rad H, Luthman H (2006)Количественный анализ локусов с высоким разрешением показывает множественные локусы предрасположенности к диабету, сопоставленные с интервалами <800 т.п.н. в видоконсервативном Niddm1i крысы GK. Генетика 174: 1565–1572.
- 17.Уэйн Л.В., Армор Дж.А., Тобин М.Д. (2009)Изменение числа копий генома, здоровье человека и болезни. Ланцет 374: 340–350.
- 18. Мегарбейн А., Равель А., Мирчер С., Стурц Ф., Граттау Ю. и др. (2009) 50-летие открытия трисомии 21: прошлое, настоящее и будущее исследований и лечения синдрома Дауна. Жене Мед 11: 611–616.
- 19. Редон Р., Исикава С., Фитч К.Р., Феук Л., Перри Г.Х. и другие. (2006) Глобальная вариация числа копий в геноме человека.Природа 444: 444–454.
- 20. Иган К.М., Шридхар С., Виглер М., Холл И.М. (2007)Рекуррентное изменение числа копий ДНК у лабораторной мыши. Нат Жене 39: 1384–1389.
- 21. Гурьев В., Саар К., Адамович Т., Верхел М., ван Хиш С.А. и др. (2008)Распределение и функциональное влияние изменения количества копий ДНК у крыс. Нат Жене 40: 538–545.
- 22. Cook EH Jr, Scherer SW (2008)Вариации числа копий, связанные с нейропсихиатрическими состояниями.Природа 455: 919–923.
- 23. Морроу Э.М., Ю С.И., Флавелл С.В., Ким Т.К., Лин И и др. (2008) Идентификация локусов и генов аутизма путем отслеживания недавних общих предков. Наука 321: 218–223.
- 24. Schaschl H, Aitman TJ, Vyse TJ (2009)Изменение количества копий в геноме человека и его влияние на аутоиммунитет. Clin Exp Immunol 156: 12–16.
- 25. Маккэрролл С.А., Альтшулер Д.М. (2007)Количество копий и исследования ассоциации заболеваний человека.Нат Жене 39: S37–42.
- 26. WTCCC (2010) Полногеномное исследование ассоциации CNV в 16 000 случаев восьми распространенных заболеваний и 3 000 общих контролей. Природа 464: 713–720.
- 27. Барретт Т., Троуп Д.Б., Уилхайт С.Е., Леду П., Руднев Д. и др. (2009) NCBI GEO: архив высокопроизводительных функциональных геномных данных. Рез. нуклеиновых кислот 37: D885–890.
- 28. Stocker R (2009)Молекулярные механизмы, лежащие в основе антиатеросклеротического и антидиабетического действия пробукола, сукцинобукола и других аналогов пробукола.Curr Opin Lipidol 20: 227–235.
- 29. Ким С.Дж., Рамеш С., Гупта Х., Ли В. (2007)Взаимодействие таурина и диабета: от участия к защите. Агенты J Biol Regul Homeost 21: 63–77.
- 30. Graubert TA, Cahan P, Edwin D, Selzer RR, Richmond TA, et al. (2007) Карта высокого разрешения вариаций числа копий сегментной ДНК в геноме мыши. PLoS Genet 3: e3.
- 31. Li Y, Agarwal P (2009) Взгляд на болезни человека и взаимосвязь между болезнями на основе путей.ПЛОС Один 4: e4346.
- 32. Teber ET, Liu JY, Ballouz S, Fatkin D, Wouters MA (2009)Сравнение автоматизированных систем предсказания генов-кандидатов с использованием генов, вовлеченных в диабет 2 типа, с помощью полногеномных ассоциативных исследований. BMC Bioinformatics 10: Приложение 1S69.
- 33. Tiffin N, Adie E, Turner F, Brunner HG, van Driel MA, et al. (2006) Вычислительная идентификация генов болезней: набор методов отдает приоритет генам-кандидатам диабета 2 типа и ожирения.Нуклеиновые кислоты Рез. 34: 3067–3081.
- 34. Беджерано Г., Фазан М., Макунин И., Стивен С., Кент В.Дж. и др. (2004) Ультраконсервативные элементы в геноме человека. Наука 304: 1321–1325.
- 35. Derti A, Roth FP, Church GM, Wu CT (2006) Ультраконсервативные элементы млекопитающих сильно истощены среди сегментных дупликаций и вариантов числа копий. Нат Жене 38: 1216–1220.
- 36. He L, Hannon GJ (2004)МикроРНК: малые РНК, играющие большую роль в регуляции генов.Нат Рев Генет 5: 522–531.
- 37. Croce CM (2009)Причины и последствия дисрегуляции микроРНК при раке. Нат Рев Генет 10: 704–714.
- 38. Tang X, Muniappan L, Tang G, Ozcan S (2009) Идентификация регулируемых глюкозой микроРНК из {бета} клеток поджелудочной железы выявляет роль miR-30d в транскрипции инсулина. РНК 15: 287–293.
- 39. Энрайт А.Дж., Джон Б., Галл У., Тушл Т., Сандер С. и др. (2003)МикроРНК-мишени у дрозофилы.Геном Биол 5: R1.
- 40. Эльбейн С.К., Дас С.К., Холлман Д.М., Ханис С.Л., Хасстедт С.Дж. (2009)Геномное сцепление и картирование примесей диабета 2 типа в афроамериканских семьях из исследовательской когорты Американской диабетической ассоциации GENNID (Генетика NIDDM). Диабет 58: 268–274.
- 41. Kroetz DL, Yook P, Costet P, Bianchi P, Pineau T (1998)Рецептор, активируемый пролифератором пероксисом альфа, контролирует печеночный адаптивный ответ индукции CYP4A на голодание и диабет.J Biol Chem 273: 31581–31589.
- 42. Алмон Р.Р., Дюбуа Д.К., Лай В., Сюэ Б., Ни Дж. и др. (2009) Анализ экспрессии генов роли печени в возникновении и развитии диабета у крыс Goto-Kakizaki. Дж. Эндокринол 200: 331–346.
- 43. Хуанг Б., Цинь В., Чжао Б., Ши Ю., Яо С. и др. (2009)Профилирование экспрессии микроРНК в модели крыс с диабетом GK. Acta Biochim Biophys Sin (Шанхай) 41: 472–477.
- 44. Ван С. Х., Цянь Р. З., Чжан В., Чен С. Ф., Джин Х. М. и др.(2009)Экспрессия микроРНК-320 в эндотелиальных клетках микрососудов миокарда и ее связь с инсулиноподобным фактором роста-1 у крыс с диабетом 2 типа. Clin Exp Pharmacol Physiol 36: 181–188.
- 45. Ловис П., Роггли Э., Лейбатт Д.Р., Гаттеско С., Ян Дж.И. и другие. (2008) Изменения в экспрессии микроРНК способствуют индуцированной жирными кислотами дисфункции бета-клеток поджелудочной железы. Диабет 57: 2728–2736.
- 46. Lu H, Buchan RJ, Cook SA (2010) МикроРНК-223 регулирует экспрессию Glut4 и метаболизм глюкозы в кардиомиоцитах.Cardiovasc Res 86: 410–420.
- 47. Клотинг Н., Бертольд С., Ковач П., Шон М.Р., Фассхауэр М. и соавт. (2009)Экспрессия микроРНК в сальниковой и подкожной жировой ткани человека. ПЛОС ОДИН 4: e4699.
- 48. Herrera BM, Lockstone HE, Taylor JM, Wills QF, Kaisaki PJ, et al. (2009) МикроРНК-125a сверхэкспрессируется в тканях-мишенях инсулина в модели спонтанного диабета 2 типа у крыс. BMC Med Genomics 2: 54.
- 49. Ортега Ф.Дж., Морено-Наваррете Дж.М., Пардо Г., Сабатер М., Хуммель М. и др.(2010) Профиль экспрессии микроРНК в подкожной жировой клетчатке человека и во время дифференцировки адипоцитов. PLoS One 5: e9022.
- 50. Допман Э.Б., Хартл Д.Л. (2007)Портрет полиморфизма числа копий у Drosophila melanogaster. Proc Natl Acad Sci U S A 104: 19920–19925.
- 51. Купер Г.М., Никерсон Д.А., Эйхлер Э.Е. (2007)Мутационное и селективное воздействие на варианты числа копий в геноме человека. Нат Жене 39: S22–29.
- 52. Смит М.Л., Мариони Дж., Хардкасл Т.Дж., Торн Н.П. (2006) snapCGH: Руководство пользователя по сегментации, нормализации и обработке данных aCGH.Биопроводник.
- 53. Neuvial P, Hupe P, Brito I, Liva S, Manie E и др. (2006) Пространственная нормализация данных массива CGH. BMC Биоинформатика 7: 264.
- 54. Smyth GK, Speed T (2003) Нормализация данных микрочипов кДНК. Методы 31: 265–273.
- 55. Смит Г.К. (2004)Линейные модели и эмпирические байесовские методы для оценки дифференциальной экспрессии в экспериментах с микрочипами. Stat Appl Genet Mol Biol 3: Статья3.
- 56.Олшен А.Б., Венкатраман Э.С., Лусито Р., Виглер М. (2004)Круговая бинарная сегментация для анализа данных о количестве копий ДНК на основе массива. Биостатистика 5: 557–572.
- 57. Awad IA, Rees CA, Hernandez-Boussard T, Ball CA, Sherlock G (2004) Caryoscope: Java-приложение с открытым исходным кодом для просмотра данных микрочипов в геномном контексте. BMC Биоинформатика 5: 151.
- 58. Двинелл М.Р., Уорти Э.А., Шимояма М., Бакир-Гунгор Б., Депонс Дж. и др. (2009) База данных генома крысы 2009: вариации, онтологии и пути.Рез. нуклеиновых кислот 37: D744–749.
- 59. Rhead B, Karolchik D, Kuhn RM, Hinrichs AS, Zweig AS, et al. (2010) База данных UCSC Genome Browser: обновление 2010 г. Nucleic Acids Res 38: D613–619.
- 60. Kent WJ (2002) BLAT – инструмент выравнивания, подобный BLAST. Геном Res 12: 656–664.
- 61. Griffiths-Jones S, Saini HK, van Dongen S, Enright AJ (2008) miRBase: инструменты для геномики микроРНК. Рез. нуклеиновых кислот 36: D154–158.
- 62.Benjamini Y, Hochberg Y (1995) Контроль частоты ложных открытий: практичный и мощный подход к множественному тестированию. Журнал Королевского статистического общества, серия B 289–300.
Вегетарианская и веганская диеты и риски тотальных и локальных переломов: результаты проспективного исследования EPIC-Oxford | BMC Medicine
Резюме результатов
В целом, веганы в этом исследовании имели более высокий риск тотальных переломов и переломов некоторых локализаций (бедренной кости, ноги, позвонка), чем мясоеды.Самые сильные ассоциации наблюдались для переломов шейки бедра, риск которых был выше у любителей рыбы, вегетарианцев и веганов. Эти различия в риске могут быть частично объяснены более низким средним ИМТ и более низким средним потреблением кальция и белка у лиц, не употребляющих мясо. Однако, поскольку различия остались, особенно у веганов, после учета этих факторов могут оказаться важными и другие неучтенные факторы.
Сравнение с предыдущими исследованиями
В нескольких предыдущих исследованиях изучалась связь вегетарианской диеты с риском переломов.В предыдущем анализе EPIC-Oxford переломов, о которых сообщали сами пациенты, с коротким последующим наблюдением, у веганов, но не у рыбоядных или вегетарианцев, риск общих переломов был на 30% (HR 1,30; 1,02, 1,66) выше, но в отличие от Согласно текущим результатам, связь полностью ослабла, когда она была ограничена участниками, которые сообщили о потреблении не менее 525 мг кальция в день [16]. Это очевидное несоответствие можно объяснить несколькими различиями между текущим и предыдущим анализом; в то время как текущий анализ включал около 4000 госпитализированных случаев в течение более чем 17 лет среднего наблюдения около 55 000 участников, предыдущее исследование включало менее 2000 случаев переломов, о которых сообщали сами, в течение 5 лет наблюдения около 35 000 участников.Учитывая разницу в методах подтверждения случая, текущий анализ менее подвержен ошибкам в отчетах и не подвержен выборочному отсеву. Также возможно, что было недостаточно мощности для обнаружения различий после стратификации по статусу потребления кальция в предыдущем анализе, в котором также не изучались локальные переломы.
Единственными другими исследованиями, в которых сообщалось о рисках переломов в зависимости от диеты, было одно небольшое проспективное исследование во Вьетнаме с участием 210 женщин (105 веганов), в котором не было обнаружено существенной разницы в частоте переломов (всего 10 случаев) между веганами и всеядными в течение 2 лет. [17] и одно проспективное исследование в Индии, в котором сообщалось о более высокой общей частоте стрессовых переломов (всего 604 случая) среди 2131 вегетарианца, чем среди 6439 новобранцев-невегетарианцев [18].Отдельно предыдущие результаты Адвентистского исследования здоровья 2, в котором большая часть вегетарианцев, показали, что участники, которые ели мясо более трех раз в неделю, имели более низкий риск переломов бедра (ОР 0,60; 0,41, 0,87), чем участники, которые ели мясо. менее одного раза в неделю [32], в то время как комбинированный анализ женщин в пери- и постменопаузе из Adventist Health Study 1 и 2 показал, что у участников, которые ели мясо более четырех раз в неделю, был более низкий риск переломов запястья (ОР 0,44; 0,23, 0). .84), чем у участников, которые никогда не ели мяса [33], но эти результаты нельзя использовать для вывода о рисках у любителей рыбы, вегетарианцев или веганов как отдельных диетических групп.
Интерпретация результатов и последствия
Более высокий наблюдаемый риск переломов у лиц, не употребляющих мясо, обычно был выше до корректировки ИМТ, что позволяет предположить, что различия в риске, вероятно, были частично обусловлены различиями в ИМТ. Вегетарианцы и веганы, как правило, имеют более низкий ИМТ, чем мясоеды [2, 8], и в предыдущих исследованиях сообщалось об обратной связи между ИМТ и некоторыми переломами, особенно переломами шейки бедра, возможно, по причинам, включая амортизацию ударной силы во время падения, повышенный уровень эстрогена. производство с повышенным ожирением или более крепкие кости из-за увеличения нагрузки [14, 34].Однако положительная связь между ИМТ и риском перелома наблюдалась для некоторых других мест, включая переломы лодыжки, возможно, в результате более высоких крутящих моментов от скручивания лодыжки у людей с более высоким ИМТ [14]. В нашем исследовании не наблюдалось существенных различий в рисках переломов лодыжек в зависимости от диеты, но точечные оценки прямо согласовывались с более низким риском у всех, кто не ел мясо, до корректировки ИМТ, и результаты могут отражать противовес между защитным эффектом от более низкого ИМТ, но более высокого риска из-за более низкого потребления питательных веществ, связанных со здоровьем костей, у тех, кто не ест мясо.
В нашем стратифицированном анализе имеются ограниченные данные о неоднородности риска переломов по категориям ИМТ. Хотя статистически значимо более высокий риск тотальных переломов и переломов бедра наблюдался только у веганов с более низким ИМТ (< 22,5 кг/м 2 ), наша интерпретация ограничена небольшим количеством случаев в каждой страте в этих анализах, особенно из-за сильной корреляции между диетической группой и ИМТ, что приводит к очень небольшому количеству веганов с более высоким ИМТ и, наоборот, сравнительно небольшому количеству мясоедов с низким ИМТ.В дополнение к ИМТ в предыдущих исследованиях сообщалось, что мышечная сила является важным фактором риска, защищающим от риска падения и последующего перелома у пожилых людей [35]. Предыдущее исследование, проведенное в Великобритании, выявило более низкую мышечную массу и силу хвата у вегетарианцев и веганов по сравнению с мясоедами [2]; поэтому необходимо дополнительно изучить возможное влияние мышечной силы и риска падения в дополнение к здоровью костей на риск переломов у вегетарианцев и веганов. Переломы в некоторых местах, особенно в области тазобедренного сустава, также могут быть в большей степени связаны с остеопорозом, чем переломы в некоторых других местах, которые с большей вероятностью могут быть результатом сильных ударов при несчастных случаях [36, 37].В этом исследовании мы не смогли дифференцировать фраг- ментальные и травматические переломы, так как отсутствовали данные о причинах переломов.
В этом и предыдущих исследованиях веганы потребляли значительно меньше кальция, чем другие диетические группы, поскольку они не потребляли молочные продукты, являющиеся основным источником диетического кальция [4, 5], в то время как вегетарианцы и веганы в среднем потребляли меньше белка [6, 7]. В организме человека 99% кальция находится в костях и зубах в виде гидроксиапатита, который в случае дефицита кальция резорбируется для поддержания метаболического баланса кальция, и, таким образом, остеопороз может возникнуть, если кальций не восстанавливается [38]. ,39,40].В недавнем метаанализе сообщалось, что увеличение потребления кальция из пищевых источников или пищевых добавок приводило к небольшому увеличению МПК [9], но данные о риске переломов были менее последовательными. Предыдущие анализы в EPIC-Oxford выявили более высокий риск переломов у женщин, но не у мужчин, при потреблении кальция ниже 525 мг/день по сравнению с более чем 1200 мг/день [41]. Недавний метаанализ как рандомизированных, так и проспективных исследований показал, что не было доказательств связи между потреблением кальция с пищей и риском переломов, но возможна слабая защитная связь между приемом добавок кальция и некоторыми переломами [10].Однако совсем недавно отдельный метаанализ показал защитный эффект от переломов комбинированных добавок витамина D и кальция, но не только добавок витамина D [11].
Что касается белка, некоторые более ранние исследования показали, что чрезмерное потребление белка приведет к увеличению метаболической кислотной нагрузки, которая впоследствии будет компенсироваться резорбцией кости и кальциурией, и, таким образом, к ухудшению здоровья костей [12, 42]. Однако более поздние экспериментальные данные показали, что высокое потребление белка также увеличивает всасывание кальция в кишечнике [43] и стимулирует выработку инсулиноподобного фактора роста (IGF)-I [44], что, в свою очередь, связано с улучшением здоровья костей [44]. 45, 46].В двух метаанализах, включавших разные исследования, сообщалось о возможном защитном эффекте повышенного потребления белка на МПК поясничного отдела позвоночника [13, 47]; в нескольких эпидемиологических исследованиях сообщалось об обратной связи между потреблением белка и риском переломов [48,49,50], хотя недавний метаанализ не обнаружил значимой связи между потреблением белка и остеопорозными переломами [51].
Более высокий риск переломов, особенно у веганов, оставался значительным после корректировки содержания кальция и белка в рационе, что позволяет предположить, что эти факторы могут лишь частично объяснять различия в рисках переломов в зависимости от диеты, и другие факторы также могут вносить свой вклад.Однако оценка потребления этих нутриентов с помощью опросников имеет существенную ошибку, и мы смогли учесть только различия в содержании кальция в рационе, но не различия в использовании добавок кальция, поскольку данные о последнем отсутствовали. Подробный анализ связи определенных пищевых продуктов, таких как мясо или молочные продукты, с риском переломов выходит за рамки настоящего исследования, но его следует изучить в дальнейших исследованиях. Будущие исследования также должны быть сосредоточены на возможном влиянии других питательных веществ или биологических маркеров на риск переломов, например, циркулирующего витамина D, витамина B 12 или IGF-I, которые могут варьироваться в зависимости от степени потребления пищи животного происхождения [52,53]. ,54].Следует также дополнительно изучить значение включения привычных пищевых привычек в дополнение к установленным параметрам для прогнозирования риска переломов в клинических условиях.
Сильные стороны и ограничения
Сильные стороны этого исследования заключались в том, что оно включало большое количество людей, не употребляющих мясо, с длительным наблюдением и изучало как тотальные, так и локализованные переломы после учета ряда искажающих факторов. Мы обновили группу диеты и соответствующие вмешивающиеся факторы, где это возможно, чтобы учесть изменения за период наблюдения.Было мало доказательств обратной причинно-следственной связи, поскольку результаты были схожими после исключения первых 5 лет наблюдения. Данные об исходах были установлены на основе больничных записей, что уменьшило количество неверных сообщений и выборочную потерю для последующего наблюдения, хотя возможным ограничением этого подхода было то, что менее серьезные переломы, не требующие госпитализации, не были бы зафиксированы.
Из других ограничений, хотя мы исключили известные случаи переломов до исходного уровня на основании больничных записей, это может не быть полным исключением, так как вопросы о предыдущем диагнозе переломов (до самых ранних доступных больничных данных) или остеопорозе не задавались при исходный уровень, а данные об использовании препаратов против остеопороза отсутствовали.Повторные измерения диеты не были доступны для всех участников, и точная дата изменения диеты во время последующего наблюдения также не была зафиксирована, но, учитывая хорошее согласие диетической группы у участников, которые проводили повторные измерения, и тот факт, что диета изменение может повлиять на риск перелома только через некоторое время, мы не ожидаем существенной неправильной классификации. Как и во всех обсервационных исследованиях, могут присутствовать остаточные искажения как диетических, так и недиетических факторов; например, роль кальция могла быть недооценена из-за ошибки измерения.Поскольку в исследование входят преимущественно белые европейцы, возможность обобщения на другие популяции или этнические группы может быть ограничена, что может быть важно, учитывая ранее наблюдаемые различия в МПК [2, 55] и риске переломов [56] в зависимости от этнической принадлежности. Мы также наблюдали лишь небольшое количество случаев во многих анализах подгрупп, и поэтому, вероятно, у нас не было достаточной мощности, чтобы надежно оценить, может ли быть какая-либо гетерогенность по этим подгруппам, включая возраст, пол, менопаузальный статус или ИМТ; поэтому необходимы дополнительные данные для подтверждения или опровержения возможных различий.В частности, поскольку когорта EPIC-Oxford состоит преимущественно из женщин (77%), дальнейшая работа должна проводиться в когортах с большей долей мужчин, чтобы изучить гетерогенность по полу и получить надежные оценки по полу.
| Мози К.П., К. Биркеланд и П.К. Дейтон. 1968. Питательное поведение астероидов и реакция их жертв на бегство в районе Пьюджет-Саунд, 49:603-619. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Dayton PK, GA Robillard and AL DeVries.1969. Якорный ледяной фундамент в проливе Мак-Мердо, Антарктида, и его биологические эффекты. Science, 163: 273-274 | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Робиллиард Г.А. и П.К. Дейтон. 1969. Заметки о биологии хенихтиевых рыб Pagetopsis macropterus из пролива Мак-Мердо, Антарктида. Антарктический журнал США, 4(6):304-306. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Dayton PK, GA Robillard and RT Paine. 1970. Зонирование бентической фауны в результате образования якорного льда в проливе Мак-Мердо, Антарктида.В Экология Антарктики под редакцией Мартина Холдгейта. Лондон: Academic Press, стр. 244-257 | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Дейтон П.К. и С. Мартин. 1971. Наблюдения за ледяными сталактитами в проливе Мак-Мердо, Антарктида. Journal of Geophysical Research, 76(6): 1595-1599 | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Dayton PK. 1971. Последствия загрязнения бентоса пролива Мак-Мердо. Антарктический журнал США, 8(3):53-56. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Dayton PK.1971. Конкуренция, возмущение и общественная организация: предоставление и последующее использование пространства в каменистом приливном сообществе. Экологическая монография, 41:351-389. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Робиллиард Г.А. и П.К. Дейтон. 1972. Новый вид платиктенового гребневика, Lyroceteis flavorpallidus , sp. nov., из пролива Мак-Мердо, Антарктида. Канадский журнал зоологии, 50:47-52 | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Дейтон П.К. и Р.Р. Хесслер.1972. Роль биологических нарушений в поддержании разнообразия в глубоководных районах. Глубоководные исследования, 19: 199-208. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Dayton PK. 1972 г. На пути к пониманию устойчивости сообщества и потенциальных эффектов обогащения бентоса в проливе Мак-Мердо, Антарктика. В: Материалы коллоквиума по проблемам сохранения Антарктиды, под редакцией Б. С. Паркера. Лоуренс, Канзас. Аллен Пресс. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Dayton PK.1973. Распространение, распространение и сохранение однолетней литоральной водоросли Postelsia palmaeformis Reprecht. Экология, 52(2):433-438. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Dayton PK. 1973. Два случая разделения ресурсов в приливном сообществе: правильный прогноз по неправильной причине. Американский натуралист, 107: 662-670. . | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Dayton PK, R Rosenthal and LC Mahen. 1973. Сообщества водорослей на Чилийском архипелаге: круиз R/V Hero 72-5.Antarctic Journal of the U.S., 8(2): 34-35 | РЕЗЮМЕ | |
| Dayton PK. Обзор жизни между приливами на Скалистых берегах Т. А. и А. Стивенсона. 1973. Science, 180: 176-177 | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Dayton PK, GA Robillard, RT Paine and LB Dayton. 1974. Биологическое приспособление в бентическом сообществе в проливе Мак-Мердо, Антарктида. Ecological Monographs, 44: 105-128 | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Dayton PK.1974. Сообщества водорослей на юге Южной Америки. Антарктический журнал США, 9: 22-23. | РЕФЕРАТ | |
| Розенталь Р.Дж., У.Д. Кларк и П.К. Дейтон. 1974. Экология и естественная история насаждения гигантских водорослей, Macrocystis pyrifera , у побережья Дель-Мар, Калифорния. Бюллетень рыболовства, 72: 672-684. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Dayton PK. 1975. Экспериментальная оценка экологического доминирования в каменистом приливно-отливном сообществе водорослей.Экологические монографии, 45(2):137-159. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Dayton PK. 1975. Экспериментальные исследования взаимодействия водорослей в сообществе водорослей, где преобладают морские выдры, на острове Амчитка, Аляска. Вестник рыболовства, 73(2): 230-237. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Dayton PK. 1975. Бентические сообщества пролива Мак-Мердо. Антарктический журнал США, X(4):136. | РЕФЕРАТ | |
| Оливер Дж.С., Д.Дж. Уотсон, Э.Ф. О’Коннер и П.К. Дейтон.1976. Бентические сообщества пролива Мак-Мердо. Antarctic Journal of the U.S., XI(2): 58-59 | РЕФЕРАТ | |
| Dayton PK and JS Oliver. 1977. Процессы, организующие контрастные бентические сообщества пролива Мак-Мердо. Антарктический журнал США, XII(4):20-21. | РЕЗЮМЕ | |
| Dayton PK, RJ Rosenthal, LC Mahen and T Antezana. 1977. Структура популяции и биология кормодобывания хищного чилийского астероида Meyenaster gelatinosus и биология побега его добычи.Морская биология, 39:361-370. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Oliver JS and PK Dayton. 1977. Антарктический мягкодонный бентос в олиготрофной и эвтрофной среде. Наука, 197:55-58. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Тегнер М.Дж. и П.К. Дейтон. 1977. Образцы морских ежей и последствия коммерческого рыболовства. Наука, 196:324-326. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Dayton PK and JS Oliver. 1978. Долгосрочные экспериментальные исследования бентоса в проливе Мак-Мердо.Antarctic Journal of the U.S., XIII(4):136-137 | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Dayton PK. 1979. Рецензия на книгу «Побережья Тихого океана: Путеводитель по экологии приливов» Т. Кэрфута. Лимнология и океанография, 24(4):792. | ОБЗОР КНИГИ | |
| Дейтон ПК. 1979. Экология: наука и религия. В: Экологические процессы в прибрежных и морских системах, под редакцией Р. Дж. Ливингстона. Нью-Йорк: Пленум Пресс, стр. 3–18. | ОБЗОР СТАТЬИ | |
| Dayton PK.1979. Наблюдения за ростом, расселением и динамикой популяций некоторых губок в проливе Мак-Мердо, Антарктида. В: Biologie des Spongiaires (Биология губки), под редакцией К. Леви и Н. Бури-Эно. Colloques Internationaux du Centre National de la Recherché Scientifique № 291, Париж, стр. 271–282. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Dayton PK. 1980. Цитата Классик. Current Contents, 32: 18. | ОБЗОРНАЯ СТАТЬЯ | |
| Dayton PK and JS Oliver.1980. Оценка экспериментального анализа структур популяций и сообществ в бентической морской среде. В: Marine Benthic Dynamics, под редакцией KR Tenore и BC Coull. Колумбия: Университет Южной Каролины Press, стр. 93-120. | ОБЗОРНАЯ СТАТЬЯ | |
| Дейтон П.К., Б.Д. Келлер и Д.А. Вен Треска. 1980. Исследования прибрежного сообщества, населенного каланом. Заключительный отчет Комиссии по морским млекопитающим США по выполнению контрактов MM6AC026 и MM1300702-9.(Национальная служба технической информации, Министерство торговли США, Спрингфилд, Вирджиния, 22161). | ОТЧЕТ ПО ИССЛЕДОВАНИЯМ | |
| Тегнер М.Дж. и П.К. Дейтон. 1981. Структура популяции, пополнение и смертность двух морских ежей Strongylocentrotus franciscanus и S. purpuratus в ламинарии. Серия «Прогресс морской экологии», 5: 255-268 | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Биркеланд С., П.К. Дейтон и Н.А. Энгстрем. 1982. Стабильная система хищничества голотурий четырьмя астероидами и их главным хищником.В: Документы конференции по иглокожим, Сидней, 1978 г., под редакцией Ф. Роу. Мемуары Австралийского музея, 16: 175–189. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Dayton PK, WA Newman and JS Oliver. 1982. Вертикальная зональность глубоководной антарктической ракушки, Bathylasma corolliforme (Hoek): экспериментальные пересадки с шельфа на мелководье. Journal of Biogeography, 9:95-109 | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Дэвис Н., Г.Р. Ван Блариком и П.К. Дейтон.1982. Искусственные сооружения на морских отложениях: воздействие на соседние бентические сообщества. Морская биология, 70:295-303 | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Dayton PK. 1982. Прибрежная экология, обзор The Shore Environment . Наука, 215:393-394. | ОБЗОР КНИГИ | |
| Дейтон ПК. 1982. Антарктида, прибрежная экология. В: Энциклопедия пляжей и прибрежной среды, под редакцией М.Л. Шварца, стр. 4-43. Издательство Хатчинсон Росс, 940 стр. | ОБЗОР СТАТЬИ | |
| Dayton PK. 1984. Процессы структурирования некоторых морских сообществ: являются ли они общими? В книге «Экологические сообщества: концептуальные вопросы и доказательства» под редакцией Д. Р. Стронга-младшего, Д. Симберлоффа, Л. Г. Абеле и А. Б. Тистла. Princeton University Press, Princeton, NJ, 613 pp. | ОБЗОРНАЯ СТАТЬЯ | |
| Dayton PK, V Currie, T Gerodette, BD Keller, R Rosenthal и DA Ven Tresca. 1984. Динамика участков и стабильность некоторых сообществ водорослей в Калифорнии.Экологические монографии, 54(3): 253-289. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Dayton PK and MJ Tegner. 1984. Катастрофические штормы, Эль-Ниньо и стабильность участка в сообществе водорослей в южной Калифорнии. Science, 224: 283-285 | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Dayton PK and MJ Tegner. 1984. Важность масштаба в экологии сообщества: пример ламинарии с наземными аналогами. В книге «Новая экология: новые подходы к интерактивным системам». стр. 457-481, под редакцией П.Прайс, С. Слободчиков и В. Гауд. John Wiley & Sons | BOOK CHAPTER | |
| Dayton PK. 1985. Структура и регулирование некоторых сообществ водорослей в Южной Америке. Экологические монографии, 55:447-468. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Dayton PK, GL Kooyman and JP Barry. 1984. Донная жизнь под толстым льдом. Antarctic Journal, 19: 128 | РЕФЕРАТ | |
| Dayton PK. 1985. Обзор экологии Антарктики.Законы RM (изд.), 2 тома, Academic Press, Орландо. Наука, 229: 157-158. . | ОБЗОР КНИГИ | |
| Барилотти, округ Колумбия, Р.Х. Макпик и П.К. Дейтон. 1985. Экспериментальные исследования воздействия коммерческого сбора ламинарии в центральной и южной Калифорнии Macrocystis pyrifera зарослей ламинарии. Calif. Fish and Game, 71(1): 4-20 | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Dayton PK. 1985. Обзор сублиторальной экологии: экология мелководного сублиторального бентоса, Р. Эрл и Д.Г. Эрвин, ред., Кларендон Пресс, Нью-Йорк. Экология, 66(1): 316-317. | ОБЗОР КНИГИ | |
| Дейтон ПК. 1985. Экология сообществ ламинарии. Аня. Преподобный Экол. и систем, 16:215-245. | ОБЗОРНАЯ СТАТЬЯ | |
| Денли Э.Дж. и П.К. Дейтон. 1985. Конкуренция среди макроводорослей. Справочник по психологическим методам, М. М. Литтлер и Д. С. Литтлер, под ред., Кембриджский унив. Press, стр. 511-530 | BOOK CHAPTER | |
| Генин А., П.К. Дейтон, П.Ф. Лонсдейл и Ф.Н. Списс.1986. Кораллы на пиках подводных гор: свидетельство ускорения течений над глубоководной топографией. Nature, 322: 59-61 | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Dayton PK. 1986. Кумулятивные воздействия в морской среде, GE Beanlands, et. др. ред., Кумулятивное воздействие на окружающую среду: двусторонняя перспектива. Канадская среда Оценка рез. Совет, Оттава и National Res. Совета, Вашингтон, округ Колумбия. стр. 79-84. | ОБЗОР ГЛАВЫ | |
| Дейтон П.К., Д. Уотсон, А. Пальмизано, Дж. П. Барри, Дж. С. Оливер и Д. Ривера.1986. Схемы распределения биотопа бентосных микроводорослей в проливе Мак-Мердо, Антарктика. Полярная биология, 6:207-213. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Тегнер М.Дж. и П.К. Дейтон. 1987. Влияние Эль-Ниньо на сообщества водорослей в южной Калифорнии. Достижения в экологических исследованиях, 17:243-279 | ОБЗОРНАЯ СТАТЬЯ | |
| Барри Дж. П. и П. К. Дейтон. 1988. Текущие закономерности в проливе Мак-Мердо, Антарктида, и их связь с местными биотическими сообществами.Полярная биология, 8:367-376. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Seymour RJ, MJ Tegner, PK Dayton и PE Parnell. 1989. Гибель гигантских водорослей, Macrocystis pyrifera , вызванная штормовой волной, в южной Калифорнии. Устье рек, прибрежная и шельфовая наука 28: 277-292. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Dayton PK, JH Carleton, AG Mackley и PW Sammarco. 1989. Модели расселения, выживания и роста устриц на Большом Барьерном рифе.Серия «Прогресс морской экологии», 54: 75-90 | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Dayton PK, RJ Seymour, PE Parnell и MJ Tegner. 1989. Необычная морская эрозия в округе Сан-Диего в результате одного шторма. Эстуарий, Наука о прибрежных зонах и шельфе, 29:151-160. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Dayton, PK. 1989. Междекадные изменения антарктической губки и ее хищников в результате океанографических климатических изменений. Наука, 245:1484-1496. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Dayton PK and MJ Tegner.1990. Дно под взволнованными водами: бентические воздействия Эль-Ниньо 1982-1984 гг. в умеренной зоне. Стр. 433-472. В: Глобальные экологические последствия Эль-Ниньо 1982-83 гг. — Южное колебание. Питер Глинн, редактор Elsevier Press, 563 стр. | ОБЗОРНАЯ СТАТЬЯ | |
| Dayton PK. 1990. Полярный бентос. Стр. 631-685. В полярной океанографии, изд. W Smith, Academic Press, 760 стр. | ОБЗОРНАЯ СТАТЬЯ | |
| Барри Дж. П. и П. К. Дейтон. 1991.Физическая неоднородность и организация морских сообществ. В книге «Экологическая гетерогенность», Дж. Коласа и С. Т. Пикетт, ред., Springer-Verlag, Нью-Йорк, стр. 269-320 | ГЛАВА КНИГИ | |
| Сделано Т., П. К. Дейтон, А. Е. Дейтон и Р. Стегер. 1991. Региональные и местные различия в восстановлении мелководных коралловых сообществ: Муреа, Французская Полинезия и центральная часть Большого Барьерного рифа. Коралловые рифы, 9: 183-192. изменение) | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Риддл М.Д., Д. Алонги, П.К. Дейтон, Дж. Хансен, Д. Клумпп.1990. Обломочные пути в лагуне кораллового рифа I. Биомасса макрофауны и оценки продукции. Морская биология, 104: 109-118. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Тегнер М.Дж. и П.К. Дейтон. 1991. Морские ежи, Эль-Ниньо и долгосрочная стабильность сообществ ламинарии в Южной Калифорнии. Серия «Прогресс морской экологии», 77:49-63 | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Dayton PK, MJ Tegner, PE Parnell, PB Edwards. 1992. Временные и пространственные закономерности нарушения и восстановления в сообществе ламинарии.Экологическая монография, 62: 421-445. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Dayton PK. 1992. Научная коммуникация — Бремя ответственности. TREE, 7(11): 392. | ПИСЬМО | |
| Хансен Дж. А., Д. В. Клумпп, Д. М. Алонги, П. К. Дейтон и М. Дж. Риддл. 1992. Обломочные пути в лагуне кораллового рифа. II. Отложение детрита, бентическая микробная биомасса и продукция. Морская биология, 113: 363-372 | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Дейтон П.К., Б.Дж. Мордида и Ф. Бэкон.1994. Полярные морские сообщества. Американский зоолог, 34(1):90-99. | ОБЗОР | |
| Dayton PK. 1994. Ландшафт сообщества: масштаб и устойчивость морских сообществ с твердым дном. стр. 289-332, глава 10 в водной экологии: масштабы, закономерности и процессы. П.С. Гиллер, А.Г. Хилдрю и Д.Г. Раффаэлли (редакторы). Блэквелл, Оксфорд. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Тегнер М.Дж., П.К. Дейтон, П.Б. Эдвардс, К.Л. Райзер, Д.Б. Чедвик, Т.А. Дин и Л. Дейшер. 1995.Последствия крупного разлива сточных вод для сообщества ламинарии: катастрофа или нарушение? Marine Environmental Research, 40: 181-224 | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Dayton PK, SF Thrush, MT Agardy and RJ Hofman. 1995. Воздействие морского рыболовства на окружающую среду. Aquatic Conservation: Marine and Freshwater Ecosystems, 5: 205-232 | VIEWPOINT | |
| Тегнер М.Дж., П.К. Дейтон, П.Б. Эдвардс и К.Л. Райзер. 1995. Кавитация морских ежей гигантских водорослей ( Macrocystis pyrifera C.Agardh) и их влияние на смертность водорослей в большом калифорнийском лесу. жур. эксп. Мар биол. and Ecol., 191: 83-99 | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Thrush SF, JE Hewitt, VJ Cummings and PK Dayton. 1995. Влияние нарушения среды обитания гребешковыми драгами на морские донные сообщества: что можно предсказать по результатам экспериментов? Мар. Экол. прогр. Ser., 129: 141-150 | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Dunton KH и PK Dayton.1995. Биология высокоширотных водорослей. В: HR Skjoldal, C Hopkins, KE Erikstad и HP Leinaas (ред.), стр. 499-507, Экология фьордов и прибрежных вод. Elsevier Science B.V. | ОБЗОРНАЯ СТАТЬЯ | |
| Knox GA. 1995. Биология Южного океана, В: Исследования полярных исследований, 1994. TREE, 10(8): 342. (Рецензия Пола К. Дейтона) | ОБЗОР КНИГИ | |
| Хьюстон М.А. 1995. Биологическое разнообразие; Сосуществование видов в меняющихся ландшафтах.Издательство Кембриджского университета, 681 стр. J. Exp. Мар биол. и экол., 191: 119-132. (Рецензия П.К. Дейтона) | ОБЗОР КНИГИ | |
| Гросс К.Л., С.Э. Пейк, Э. Аллен, К. Бледсо, Р. Колвелл, П.К. и С. Стаффорд. 1995. Заключительный отчет Комитета Экологического общества Америки о будущем долгосрочных экологических данных, стр. 1-122 (FLED). Том I: Текст отчета. БЕЖАЛ. | ОТЧЕТ | |
| Комитет Национальной академии наук по биологическому разнообразию морских систем, понимание морского биоразнообразия.1995. Вашингтон, округ Колумбия. National Academy Press, 75 стр. Coastal Management, 23: 337-340. (Рецензия П.К. Дейтона) | ОБЗОР КНИГИ | |
| Тегнер М.Дж., П.К. Дейтон, П.Б. Эдвардс и К.Л. Райзер. 1996. Имеются ли доказательства долгосрочных климатических изменений в водорослевых лесах Южной Калифорнии? Отчеты CalCOFI, 37, 111–126. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Тегнер М.Дж., Л.В. Баш и П.К. Дейтон. 1996. Почти исчезновение эксплуатируемого морского беспозвоночного. TREE, 11: 278-280 | RESEARCH ARTICLE | |
| Mangel M и еще 40 (PK Dayton — двенадцатый автор).1996. Принципы сохранения диких живых ресурсов. Экологические приложения, 6(2): 338-362. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Hobday AJ, L Haury and PK Dayton. 1997. Функция девственной плевы человека. Medical Hypotheses, 49(2): 171-173 | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Кауфман Л. и П.К. Дейтон. 1997. Воздействие добычи морских ресурсов на экосистемные услуги и устойчивость. В Nature’s Services, Социальная зависимость от природных экосистем, GC Daily (изд.), стр. 275-293. [Island Press] | BOOK CHAPTER | |
| Лежандр П., С.Ф. Траш, В.Дж. Каммингс, П.К. Дейтон, Дж. Грант, Дж.Э. Хьюитт, А.Х. Хайнс, Б.Х. Макардл, Р.Д. Уилкинсон. 1997. Пространственная структура двустворчатых моллюсков на песчаной отмели: процессы масштабирования и образования. Дж. Эксп. Mar. Bio., 216(1-2): 99-128 | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Hines AH, RB Whitlatch, SF Thrush, JE Hewitt, VJ Cummings, PK Dayton и P Legendre.1997. Нелинейная реакция крупного морского хищника в поисках пищи на бентосную добычу: орляковые ямки и двустворчатые моллюски на песчаной отмели Новой Зеландии J. Exp. Mar. Bio., 216(1-2): 191-210 | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Thrush SF и 19 других (с П.К. Дейтоном в качестве пятого автора). 1997. Песчаная среда обитания: от экспериментов к выводам. жур. Эксп. Мар биол. и экол., 216: 1-9. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Schneider DC, R Walters, S Thrush and P Dayton.1997. Увеличение масштабов экологических экспериментов: изменение плотности подвижного двустворчатого моллюска Macomona liliana. жур. Эксп. Мар биол. and Ecol., 216:129-152 | RESEARCH ARTICLE | |
| Thrush SF и еще 14 (с П.К. Дейтоном в качестве третьего автора). 1997. Сопоставление результатов мелкомасштабных экспериментов по манипулированию плотностью с более крупными моделями — пример взаимодействия взрослых и молодых двустворчатых моллюсков. жур. Эксп. Мар биол. и экол., 216:153-169. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Thrush SF и еще 12 (с П.К. Дейтоном в качестве пятого автора).1997. Переход от экспериментов к сложным экологическим системам: куда двигаться дальше? жур. Эксп. Мар биол. и экол., 216:243-254. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Dayton PK, KW England и EA Robson. 1997. Необычный морской анемон, Dactylanthus antarcticus (Clubb, 1908) (Отряд Ptychodactiaria), о горгонариях в чилийских фьордах, 135–142 стр. В материалах 6-й Международной конференции по кишечнополостной биологии. Левенхорст, Нордвейкерхаут, Нидерланды, 16–21 июля 1995 г. / JC den Hartog [ред.] Лейден: Национальный музей естественной истории | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Тегнер М.Дж. и П.К. Дейтон. 1997. Изменение исходных условий и проблема снижения ожиданий при прибрежном рыболовстве. проц. конф. амер. соц. Civil Engrs., Калифорния и Мировой океан ’97, Сан-Диего, Калифорния, 24–27 марта, стр. 119–128. | ПРОЦЕССЫ | |
| Тегнер М.Дж., П.К. Дейтон, П.Б. Эдвардс и К.Л. Райзер. 1997. Крупномасштабные низкочастотные океанографические воздействия на сукцессию ламинарии: рассказ о двух когортах.Мар. Экол. прогр. сер. 146: 117-134. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Dayton PK. 1998. Перекладывание бремени доказывания в управлении рыболовством. Наука 279: 821-822. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Веттер Э.В. и П.К. Дейтон. 1998. Сообщества макрофауны внутри и вокруг богатой детритом системы подводных каньонов. Deep-Sea Research Part II, 45(1-3): 25-54 | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Thrush SF, JE Hewitt, VJ Cummings, PK Dayton, M Cryer, SJ Turner, GA Funnell, RG Budd, Си Джей Милберн и М. Р. Уилкинсон.1998. Нарушение морской бентической среды обитания в результате коммерческого рыболовства: воздействие в масштабах промысла. Ecological Applications, 8(3): 866-879 | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Dayton PK, MJ Tegner, PB Edwards and KL Riser. 1998. Скользящие базовые линии, призраки и снижение ожиданий в сообществах ламинарии. Экологические приложения, 8(2): 309-322. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Дейтон П.К., М.Дж. Тегнер, П.Б. Эдвардс и К.Л. Райзер. 1999.Временные и пространственные масштабы демографии водорослей: роль океанографического климата. Ecological Monographs, 69(2): 219-250 | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Wilder JR, MJ Tegner and PK Dayton. 1999. Сохранение морского биоразнообразия. Вопросы науки и техники, XV (3): 57-64. | ОБЗОР | |
| Тегнер М.Дж. и П.К. Дейтон. 1999. Воздействие рыболовства на экосистему. Тенденции в экологии и эволюции, 14:261-262. | ОБЗОР | |
| Мюррей С.Н., Р.Ф. Амброуз, Дж.А. Бонсак, Л.В. Ботсфорд, М.Х. Карр, Г.Э. Дэвис, П.К. Дейтон, Д. Готшал, Д.Р. Гандерсон, М.А. Дж. К. Огден, Дж. Рафгарден, Р. М. Старр, М. Дж. Тегнер и М. М. Йоклавич.1999. Сети запретных заповедников: поддержание популяций рыбных промыслов и морских экосистем. Рыболовство, 24:11, 11-25. | ОБЗОР | |
| Веттер Э.В. и П.К. Дейтон. 1999. Органическое обогащение детритом макрофитов и модели численности популяций мегафауны в подводных каньонах. Серия 186: 137-148 | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Флухарти, Дэвид, Пит Апарисио, Кристин Блэкберн, Джордж Бёлерт, Фелиция Коулман, Филип Конклинг, Роберт Костанца, Пол Дейтон, Роберт Фрэнсис, Дойл Ханан, Кен Хинман, Эдвард Худ, Джеймс Китчелл, Рич Лэнгтон, Джейн Любченко, Марк Мангел, Рассел Нельсон, Виктория О’Коннелл, Майкл Орбах, Майкл Сиссенвайн.1999. Экосистемное управление рыболовством: отчет Консультативной группы по экосистемным принципам Конгрессу. 54 стр. | ТЕХНИЧЕСКИЙ ОТЧЕТ | |
| Меффе Г.К., В.Ф. Перрин и П.К. Дейтон. 1999. Сохранение морских млекопитающих: Руководящие принципы и их реализация. В области сохранения и управления морскими млекопитающими. JH Twiss Jr и RR Reeves, ред., 437-454. [Smithsonian Institution Press] | BOOK CHAPTER | |
| Талли Т.С., П.К. Дейтон и С.Э. Ибарра-Обандо.2000. Сообщества макрофауны приливных равнин и связанные с ними среды в устьях рек Южной Калифорнии и Северной Нижней Калифорнии, Мексика. Эстуарии, 23:1, 97-114. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Dayton PK, E Sala, MJ Tegner and S Thrush. 2000. Охраняемые морские территории: парки, базовые линии и улучшение рыболовства. Bulletin of Marine Science, 66(3):617-634 | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Тегнер М.Дж. и П.К. Дейтон. 2000. Экосистемные последствия рыболовства в сообществах ламинарии.Журнал морских наук ICES, 57: 579-589. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Хайренбах К.Д., К.А. Форни и П.К. Дейтон. 2000. Охраняемые районы моря и управление океаническими бассейнами. Сохранение водных ресурсов: морские и пресноводные экосистемы, 10: 437-458. | ТОЧКА ЗРЕНИЯ | |
| Мур С.Э., Д.П. ДеМастер и П.К. Дейтон. 2000. Выбор места обитания китообразных в Арктике Аляски летом и осенью. Арктика, 53:4, 432-447. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Фогарти М.Дж., Дж.А. Бонсак и П.К. Дейтон.2000. Морские заповедники и управление ресурсами. В морях тысячелетия: экологическая оценка. Том III: Глобальные проблемы и процессы. CRC Sheppard, изд., 375-392 [Пергамон]. | КНИГА ГЛАВА | |
| Уитман Дж. Д. и П. К. Дейтон. 2001. Скалистые сублиторальные сообщества. В экологии морского сообщества. MD Bertness, SD Gaines and ME Hay, eds., 339-366 [Sinauer Associates, Inc.]. | КНИГА ГЛАВА | |
| Национальный исследовательский совет. 2001.Морские охраняемые районы: инструменты для поддержания экосистем океана. (П. К. Дейтон — третий из 13 авторов). Издательство Национальной академии наук. Вашингтон. 272 стр. | КНИГА | |
| Герродетт Т., П.К. Дейтон, С. Мачинко и М. Фогарти. 2002. Предупредительное управление морским рыболовством: выход за рамки бремени доказывания. Бюллетень морской науки, 70 (2): 657-668. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Дейтон П.К. и Энрик Сала. 2001. Естествознание: чувство чуда, творчество и прогресс в экологии.Scientia Marina, 65 (Приложение 2): 199-206. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Thrush S and PK Dayton. 2002. Нарушение морских бентических местообитаний в результате траления и дноуглубительных работ: последствия для морского биоразнообразия. Аня. Преподобный Экол. Сист. 33: 449-473. | ОБЗОР | |
| Сала Э., О. Абурто-Оропеза, Г. Паредес, И. Парра, Дж. К. Баррера и П. К. Дейтон. 2002. Общая модель проектирования сетей морских заповедников. Наука 298: 1991-1993. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Dayton PK, S Thrush and FC Coleman.2002. Экологические последствия рыболовства в морских экосистемах США. Комиссия Pew Oceans, Арлингтон, Вирджиния. 45 р . | ОБЗОР | |
| Грэм М.Х. и П.К. Дейтон. 2002. Об эволюции экологических идей: парадигмы и научный прогресс. Экология 83(6): 1481-1489. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Graham MH, PK Dayton и MA Hixon. 2002. Парадигмы экологии: прошлое, настоящее и будущее. Экология 83(6): 1479-1480. | ОБЗОР | |
| Кинлан Б.П., М.Х. Грэм, Э. Сала и П.К. Дейтон.2003. Задержка развития эмбриональных спорофитов гигантских водорослей ( Macrocystis pyrifera , Phaeophyceae): механизм замедленного рекрутирования многолетних водорослей? Дж. Фикол. 39: 47-57. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Сала Э., О. Абурто-Оропеза, Г. Паредес, И. Парра, Дж. К. Баррера и П. К. Дейтон. 2003. Ответ на «Моделирование охраняемых морских территорий». Наука 301: 47-49. | ПИСЬМО | |
| Роджерс-Беннетт Л., П.Л. Хаакер, Т.О. Хафф и П.К. Дейтон.2002. Оценка исходной численности морских ушек в Калифорнии для восстановления. CalCOFI Rep., Vol 43:97-111 | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Graham MH, PK Dayton and JM Erlandson. 2003. Ледниковые периоды и экологические переходы на побережьях умеренного пояса. Тенденции в экол. и Эвол. 18(1): 33-40 | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Dayton PK. 2003. Важность естественных наук для сохранения. American Naturalist 162(1):1-13 | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Агарди Т., П. Бриджуотер, М.П. Кросби, Дж. Дэй, П.К. По.2003. Опасные цели? Нерешенные вопросы и идеологические конфликты вокруг морских охраняемых территорий. Сохранение водных ресурсов: морские и пресноводные экосистемы, 13:353-367. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Пикич Э.К., К. Сантора, Э.А. Бэбкок, А. Бакун, Р. Бонфил, Д.О. Коновер, П. Дейтон, П. Дукакис, Д. Флухарти, Б. Хенеман, Э.Д. , Дж. Поуп, К. Дж. Сейнсбери. Экосистемное управление рыболовством. Science 305:346-347 | LETTER | |
| Dayton PK.2004 г. Рецензия на книгу «Большие морские экосистемы мира: тенденции эксплуатации, защиты и исследований». Г. Хемпель, К. Шерман (ред.), 2003 г. Журнал экспериментальной морской биологии и экологии 313: 211-21 | ОБЗОР КНИГИ | |
| Вилчис Л.И., М.Дж. Тегнер, М.Д. Мур, К.С. Фридман, К.Л. Райзер, Т.Т. Роббинс и П.К. 480. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Биркеланд С. и П.К. Дейтон.2005. Важность ухода от крупных рыб в управлении рыболовством. Trends in Ecology and Evolution 20(7): 356-358 | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Parnell PE, CE Lennert-Cody, L Geelan, LD Stanley and PK Dayton. 2005 Эффективность небольшого морского заповедника в южной Калифорнии. Серия «Прогресс морской экологии», 296:39-52. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Parnell PE, PK Dayton, CE Lennert-Cody, LL Rasmussen и JJ Leichter. 2006. Дизайн морского заповедника: оптимальный размер, мелкомасштабные среды обитания, родство видов, разнообразие и микроклимат океана.Экологические приложения. 16 (3): 945-962. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Thrush S, PK Dayton, R Cattaneo-Vietti, M Chiantore, V Cummings, N Andrew, I Hawes, S Kim, R Kvitek, AM Schwarz. 2006. Широкомасштабные факторы, влияющие на биоразнообразие прибрежных донных сообществ моря Росса. Deep-Sea Research II 53: 959-971 | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Гили Дж.М., В.Е. Арнц, А. Паланкес, К. Орехас, А. Кларк, П.К. .2006. Уникальная совокупность эпибентосных сидячих взвесеядных с архаичными чертами в высокой Антарктике. Deep-Sea Research II 53:1029-1053 | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Грей Дж. С., П. Дейтон, С. Траш и М. Дж. Кайзер. 2006. О влиянии траления, бентоса и схемы отбора проб. Бюллетень о загрязнении морской среды 52:840-843. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Hewitt JE, SF Thrush, PK Dayton and E Bonsdorff. 2007. Влияние пространственной и временной неоднородности на разработку и анализ эмпирических исследований систем, зависящих от масштаба.Американский натуралист, 169:398-408. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Parnell PE, PK Dayton и F Margiotta. 2007. Пространственные и временные закономерности ловли омаров ловушками: обзор промысловых усилий и структуры среды обитания. Бык. Академик Южной Калифорнии. науч. 106 (1): 27-37. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Steneck RS, RH Bustamante, PK Dayton, GP Jones and AJ Hobday. 2008. Текущее состояние и будущие тенденции в экосистемах ламинарии. Глава 15, в водных экосистемах , изд.Н. Полунин. Издательство Кембриджского университета. | ГЛАВА КНИГИ | |
| П. Дж. Остер, Р. Фуджита, С. Р. Келлерт, Дж. Авис, К. Кампанья, Б. Кукер, П. К. Дейтон, Б. Хенеман, Р. Кенчингтон, Дж. Стоун, Г. Н. ди Скиара и П. Глинн. 2008. Развитие этики океана: наука, польза, эстетика, личные интересы и разные способы познания. Биология сохранения , 23(1): 233-235. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Hewitt JE, SF Thrush and PK Dayton. 2008.Изменчивость среды обитания, видовое разнообразие и экологическое функционирование в морской системе. Журнал экспериментальной морской биологии и экологии 366: 116-122. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Parnell PE, AK Groce, TD Stebbins and PK Dayton. 2008. Различение источников загрязнения рыб ПХБ на прибрежном шельфе Сан-Диего, Калифорния (США). Бюллетень о загрязнении морской среды 56: 1992-2002 | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Брайе Т.Дж., Дж.М. Эрландсон, Т.С. Рик, П.К. Дейтон и М.Б.А. Хэтч.2009. Рыболовство от прошлого к настоящему: преемственность и устойчивость промысла красных морских ушек на Нормандских островах, Калифорния. Экологические приложения, 19(4): 906-919. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Хофман Р. Дж., Дж. Э. Рейнольдс III, П. К. Дейтон, В. Алексадер, Д. Д. Бонесс и Т. Дж. Раген. 2009. Джон Рассел Твисс-младший (1938-2009) Наука о морских млекопитающих 25(4):1000-1003 | ПАМЯТИ | |
| Левин Л.А. и П.К. Дейтон. 2009. Экологическая теория и континентальные окраины: где мелководье встречается с глубиной.TREE 24:606-617 | ОБЗОР | |
| Thrush SF, JE Hewitt, PK Dayton, G Coco, AM Lohrer, A Norkko, J Norkko и M Chiantore. 2009. Прогнозирование пределов устойчивости: интеграция эмпирических исследований с теорией. Труды Королевского общества B, 276: 3209-3217. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Дрозд С.Ф. и П.К. Дейтон. 2010. Какой вклад экология может внести в управление на основе экосистем? Ежегодный обзор морской науки. 2: 419-41 | ОБЗОР | |
| Parnell PE, EF Miller, CE Lennert-Cody, PK Dayton, ML Carter, TD Stebbins.2010. Реакция гигантских водорослей (Macrocystis pyrifera) в южной Калифорнии на низкочастотное воздействие на климат. Limnology and Oceanography 55:2686-2702 | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Parnell PE, PK Dayton, RA Fisher, CC Loarie and RD Darrow. 2010. Пространственные модели промыслового усилия у Сан-Диего: последствия для зонального управления и функционирования экосистемы. Экологические приложения, 20:2203-2222. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Jacquet J, D Pauly, D Ainley, S Holt, PK Dayton и JBC Jackson.2010. Управление морепродуктами в условиях кризиса. Природа, 467:28-29. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Сала Э. и П.К. Дейтон. 2011. Прогнозирование сильного воздействия на сообщества с использованием экспериментальных оценок силы взаимодействия на душу населения: донные травоядные и пополнение гигантских водорослей. Морская экология, 32:300-312. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Konotchick T, PE Parnell, PK Dayton и JJ Leichter. 2012. Вертикальное распределение воздействия питательных веществ на Macrocystis pyrifera в южной Калифорнии.Эстуарий, Наука о прибрежных зонах и шельфе, 106:85-92. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Christian C, D Ainley, M Bailey, et al. 2013а. Не «лучший экологический выбор морепродуктов»: ответ Гутьерресу и Агнью (2013). Биологическая консервация 165, 214-215. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Christian C, D Ainley, M Bailey, et al. 2013б. Обзор официальных возражений против сертификации рыболовства Морским попечительским советом. Биологическая консервация 161, 10-17. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Dayton PK, S Kim, SC Jarrell, JS Oliver, K Hammerstrom, JL Fisher, K O’Connor, JS Barber, G Robillard, J Barry, AR Thurber, K Conlan. 2013. Набор, рост и смертность антарктической гексактинеллидной губки Anoxycalyx joubini. PLoS One 8 (2), e56939. DOI: 10.1371/journal.pone.0056939. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Gocke C, D Janussen, HM Reiswig, SC Jarrell, PK Dayton, 2015. Rossella podagrosa Kirkpatrick, 1907 – В конце концов, действительный вид.Зоотакса. 4021(1), 169-177. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Abrams PA, D. Ainley, LK Blight, PK Dayton, JT Eastman and JL Jacquet. 2016. Необходимые элементы предупредительного управления: последствия для антарктического клыкача Fish and Fisheries, 17, 1152-1174. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Дейтон П.К., К. Хаммерстром, С.К. Джаррелл, С. Ким, В. Нордхаузен, Д.Дж. Осборн, С.Ф.Антарктида. Антарктическая наука. DOI: 10.1017/S0954102016000171. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Dayton PK, Jarrell S, Kim S, Thrush S, Hammerstrom K, Slattery M and Parnell E. 2016. Неожиданное эпизодическое пополнение и рост антарктических губок: влияние на экологическую устойчивость. Журнал экспериментальной морской биологии и экологии. 482:38-55. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Parnell PE, JT Fumo, CE Lennert-Cody, SC Schroeter и PK Dayton.2017. Поведение морского ежа в лесу водорослей в Южной Калифорнии: еда, страх, поведенческие ниши и расширение индивидуального поведения. Журнал исследований моллюсков, 36 (2): 529-543. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Dayton PK. 2018. Миксина в заливе Салмон, пролив Мак-Мердо? Антарктическая наука. 30(4):243-244. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Dayton PK. 2018. Долгосрочное сохранение древесины на морском дне в проливе Мак-Мердо, Антарктида. Антарктическая наука.30(6), 355-356 | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Brooks C, D Ainley, P Abrams, PK Dayton, R Hofman, J Jacquet и DB Siniff. 2018. Управляйте рыболовством с большей осторожностью в Южном океане Для защиты морских экосистем вокруг Антарктики перед лицом изменения климата необходимы гибкие правила, мониторинг и запреты на рыболовство. Природа Том. 558: 177-180. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Dayton PK, SC Jarrell, S Kim, SF Thrush, K Hammerstrom и JJ Leichter.2019. Реакция бентоса на изменение режима Антарктики: размер пищевых частиц и биология пополнения. Экологические приложения. 29(1):1-20 | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ | |
| Dayton PK, JS Oliver, SF Thrush и K Hammerstrom. 2019. Бактерии защищают падаль от падальщиков. Антарктическая наука, 31(1), 13-15. | ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ |
Биомаркеры РНК крови при продромальном синдроме PARK4 и расстройстве поведения во время сна с быстрыми движениями глаз показывают роль потери комплексина 1 в риске развития болезни Паркинсона | Модели и механизмы заболеваний
После ночного голодания у людей были взяты образцы цельной крови из семейства PARK4 (после информированного письменного согласия и с одобрения Комиссии по этике Университета Богазичи в Стамбуле) в пробирки PAXgene (Qiagen) и ЭДТА.Также после ночного голодания образцы цельной крови были собраны в пробирки PAXgene у когорты RBD, состоящей из 46 больных и 19 здоровых родственников (лица с нарколепсией были исключены), после письменного информированного согласия и с одобрения Комиссии по этике Марбургского университета. Образцы от пострадавших и контрольных людей обрабатывали параллельно, и после 2-часовой инкубации пробирок PAXgene при комнатной температуре их хранили при -80 °C до экстракции РНК после дополнительной инкубации в течение ночи при комнатной температуре с использованием соответствующего набора PAXgene. .Выделенную РНК очищали с помощью ДНКазы-I степени амплификации (Invitrogen) и преобразовывали в кДНК с помощью обратной транскриптазы SuperScript III (Invitrogen). Качество и количество кДНК определяли с помощью спектрофотометрии и показателей целостности РНК (Agilent Bioanalyzer). Количественный ОТ-ПЦР проводили с анализах TaqMan (Applied Biosystems) Hs01018439_s1 ( GPRIN3 ), Hs00201182_m1 ( MMRN1 ), Hs00411197_m1 ( LRRK2 ), Hs01084510_m1 ( GIGYF2 ), Hs00223032_m1 ( ATP13A ), Hs01026447_m1 ( ATXN3 ), Hs00697109_m1 ( DJ-1 ), Hs00260868_m1 ( PINK1 ), Hs00832556_sH ( ST13 ), Hs01038318_m1 ( PARK2 ), Hs00201825_m1 ( FBX07 ), Hs00188233_m1 ( UCHL1 ), Hs00164683_m1 ( GBA ), Hs00234883_m1 ( HtrA2 ), Hs00185926_m1 ( PLA2G6 ), Hs00608185_m1 ( НОЖДБ ), Hs00705917_s1 ( YWHAG ), Hs01103386_m1 ( SNCA ), HS00793604_m1 ( YWHAB ), HS00356749_g1 ( YWHAE ), HS00362510_M1 ( CPLX1 ), HS00277212_M1 ( GZMH ), HS00959010_M1 ( SPP1 ), HS00272126_M1 ( PLTP ), HS00230853_M1 ( HNF4A ) и HS0 07_g1 ( ПТВР1 ).После исследования различных загрузки (например, GAPDH , ACTN , RPL13A , TBP , TBP , SDH и YWHA2 ) против SNCA выражение в цельной крови, HS999999910_M1 ( TBP ) наблюдалось показывают наименьшую дисперсию и поэтому были включены в качестве домашнего контроля для каждого теста экспрессии. Всего в каждой реакции использовали 25 нг кДНК. Условия ПЦР: 2 мин при 50°C, 10 мин при 95°C, 40 циклов по 15 с при 95°C и 1 мин при 60°C.Изменения экспрессии генов анализировали по методу 2 -ΔΔCt (Livak, Schmittgen, 2001).
.