
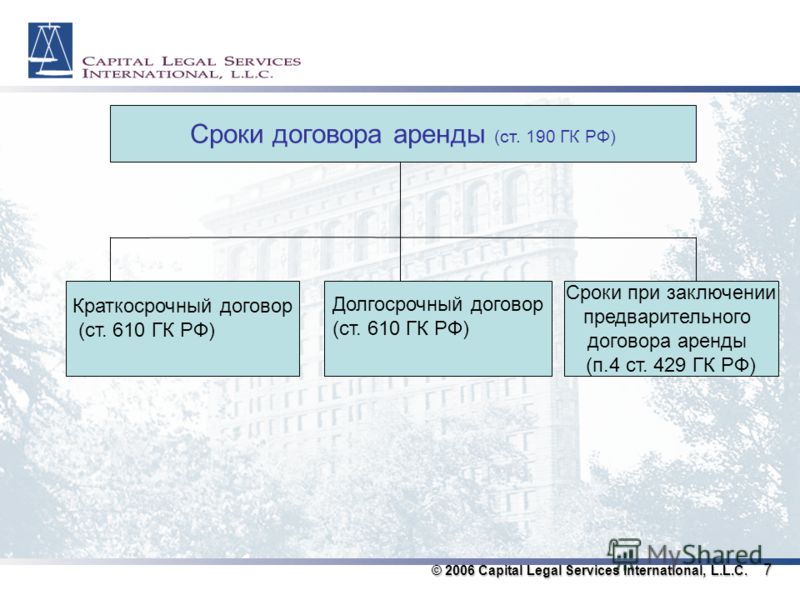
Статья 190 ГК РФ. Определение срока
Статья 190 ГК РФ. Определение срокаАктуально на:
22 января 2022 г.
Гражданский кодекс, N 51-ФЗ | ст. 190 ГК РФ
Установленный законом, иными правовыми актами, сделкой или назначаемый судом срок определяется календарной датой или истечением периода времени, который исчисляется годами, месяцами, неделями, днями или часами.
Срок может определяться также указанием на событие, которое должно неизбежно наступить.
- URL
- HTML
- BB-код
- Текст
URL документа [скопировать]
<a href=»»></a>
HTML-код ссылки для вставки на страницу сайта [скопировать]
[url=][/url]
BB-код ссылки для форумов и блогов [скопировать]
—
в виде обычного текста для соцсетей и пр. [скопировать]
[скопировать]
Скачать документ в формате
Судебная практика по статье 190 ГК РФ:
- Решение Верховного суда: Определение N ВАС-18029/12, Коллегия по гражданским правоотношениям, надзор
Суд установил: решением от 06.04.2012, оставленным без изменения постановлениями апелляционной и кассационной инстанций, иск удовлетворен. Руководствуясь статьями 190, 309, 310, 539 – 548 Гражданского кодекса Российской Федерации, договором купли-продажи тепловой энергии от 22.

- Решение Верховного суда: Определение N 78-КГ17-67, Судебная коллегия по гражданским делам, кассация
При этом передача объекта должна быть произведена не ранее получения застройщиком разрешения на ввод в эксплуатацию многоквартирного дома или иного объекта недвижимости и не позднее установленного договором срока. В соответствии со статьей 190 Гражданского кодекса Российской Федерации установленный законом, иными правовыми актами, сделкой или назначаемый судом срок определяется календарной датой или истечением периода времени, который исчисляется годами, месяцами, неделями, днями или часами…
- Решение Верховного суда: Определение N ВАС-15808/12, Коллегия по гражданским правоотношениям, надзор
Утверждение заявителя о согласовании сторонами всех существенных условий договора и необоснованном выводе судебных инстанций о нарушении требований статьи 190 Гражданского кодекса Российской Федерации о сроке осуществления мероприятий по технологическому присоединению со ссылкой на судебные акты по другим арбитражным делам, неосновательно, поскольку не опровергает выводы судебных инстанций о невозможности считать срок установленным.
..
 ..
..Постоянная ссылка на документ
- URL
- HTML
- BB-код
- Текст
URL документа [скопировать]
<a href=»»></a>
HTML-код ссылки для вставки на страницу сайта [скопировать]
[url=][/url]BB-код ссылки для форумов и блогов [скопировать]
—
в виде обычного текста для соцсетей и пр. [скопировать]
[скопировать]
Скачать документ в формате
Составить подборку
Анализ текста
Идет загрузка…
Ст. 190 ГК РФ. Определение срока
Установленный законом, иными правовыми актами, сделкой или назначаемый судом срок определяется календарной датой или истечением периода времени, который исчисляется годами, месяцами, неделями, днями или часами.
Срок может определяться также указанием на событие, которое должно неизбежно наступить.
См. все связанные документы >>>
Под сроками понимаются моменты или отрезки времени, истечение или наступление которых влечет возникновение, изменение или прекращение гражданских правоотношений. Сроки относят к событиям, поскольку течение времени не зависит от воли человека, однако установление и определение длительности срока осуществляется по воле человека.
Сроки относят к событиям, поскольку течение времени не зависит от воли человека, однако установление и определение длительности срока осуществляется по воле человека.
Сроки классифицируются по различным критериям. По правовым последствиям сроки делятся на правообразующие, правоизменяющие, правопрекращающие. В зависимости от того, могут ли сроки быть изменены по соглашению сторон, они делятся на императивные и диспозитивные. Различают также сроки абсолютно определенные (указывающие на точный период времени или момент, с которым связываются определенные последствия) и относительно определенные, которые характеризуются меньшей точностью, однако также связаны с определенным моментом или периодом времени. Неопределенные сроки имеют место тогда, когда законом или договором вообще не указан временной ориентир, однако предполагается, что срок ограничен временем.
Сроки могут устанавливаться законом (такие сроки нельзя изменить соглашением сторон, например срок приобретательной давности) и договором. Важное значение имеет деление сроков на сроки осуществления гражданских прав, сроки исполнения обязанностей и сроки защиты гражданских прав. К срокам осуществления гражданских прав относятся сроки, в течение которых управомоченный субъект может сам реализовать принадлежащее ему право либо потребовать совершения определенных действий от обязанного лица. В свою очередь, сроки осуществления гражданских прав делятся на сроки существования гражданских прав, пресекательные сроки, претензионные сроки, сроки годности и т.д.
Важное значение имеет деление сроков на сроки осуществления гражданских прав, сроки исполнения обязанностей и сроки защиты гражданских прав. К срокам осуществления гражданских прав относятся сроки, в течение которых управомоченный субъект может сам реализовать принадлежащее ему право либо потребовать совершения определенных действий от обязанного лица. В свою очередь, сроки осуществления гражданских прав делятся на сроки существования гражданских прав, пресекательные сроки, претензионные сроки, сроки годности и т.д.
К особой категории относятся пресекательные сроки. Суть их заключается в том, что они устанавливают пределы существования субъективных гражданских прав и предоставляют управомоченным лицам строго определенное время для их реализации под угрозой прекращения в случае его неосуществления или ненадлежащего осуществления.
Согласно комментируемой статье срок может определяться тремя способами: календарной датой, истечением периода времени, указанием на событие, которое неизбежно должно наступить (например, совершеннолетие).
Сроки, определяемые периодами времени, могут исчисляться любыми отрезками времени, однако чаще всего исчисляются годами, месяцами, неделями, днями. Могут применяться также такие единицы измерения времени, как полгода (п. 1 ст. 192 ГК), квартал (п. 2 ст. 192 ГК), полмесяца (п. 3 ст. 192 ГК). Сроки могут также измеряться часами, минутами или даже секундами.
Указание на событие, о котором идет речь в комментируемой статье, предполагает, что это событие неизбежно наступит. Поэтому срок не может устанавливаться, например, указанием на момент исполнения обязанной стороной своей обязанности по заключенному договору.
Верховный Суд РФ в Определении от 10 ноября 2015 г. N 80-КГ15-18 указал следующее: «Установленный законом, иными правовыми актами, сделкой или назначаемый судом срок определяется календарной датой или истечением периода времени, который исчисляется годами, месяцами, неделями, днями или часами. Срок может определяться также указанием на событие, которое должно неизбежно наступить.
Удовлетворяя требования к поручителям по настоящему делу, суд апелляционной инстанции сослался на то, что срок поручительства по договорам с юридическими лицами определен выполнением заемщиком своих обязанностей, а по договорам с физическими лицами календарной датой — 10 марта 2013 г.
Между тем выполнение заемщиком своих обязательств не является событием, которое должно с неизбежностью наступить, а следовательно, не может быть условием о сроке поручительства по смыслу статьи 190 Гражданского кодекса Российской Федерации».
Кредитные потребительские кооперативы | Банк России
Согласно положениям пункта 4 статьи 65.3 Гражданского кодекса Российской Федерации (далее — ГК РФ) в корпорации может быть образован в случаях, в частности, предусмотренных другим законом, коллегиальный орган управления (наблюдательный или иной совет), контролирующий деятельность исполнительных органов корпорации и выполняющий иные функции, возложенные на него законом или уставом корпорации. Также члены коллегиального органа управления корпорации имеют право получать информацию о деятельности корпорации и знакомиться с ее бухгалтерской и иной документацией.
Также члены коллегиального органа управления корпорации имеют право получать информацию о деятельности корпорации и знакомиться с ее бухгалтерской и иной документацией.
Одновременно в соответствии с частью 1 статьи 23 Федерального закона от 18.07.2009 № 190-ФЗ «О кредитной кооперации» (далее — Закон № 190-ФЗ) контрольно-ревизионный орган (наблюдательный совет, ревизионная комиссия или ревизор) осуществляет контроль за деятельностью КПК и его органов, а также осуществляет иные функции, предусмотренные уставом КПК. Кроме того, согласно положениям частей 3 и 4 статьи 23 Закона № 190-ФЗ контрольно-ревизионный орган обязан проводить проверку годовой бухгалтерской (финансовой) отчетности КПК до ее утверждения общим собранием членов КПК (пайщиков) и имеет право на получение от органов КПК любой информации о деятельности КПК.
Таким образом, контрольно-ревизионный орган (наблюдательный совет, ревизионная комиссия), учитывая его функционал, права и обязанности, является коллегиальным органом управления КПК (информация о его членах, являющихся аффилированными лицами, отражается по коду основания аффилированности А1-1, установленному Указанием Банка России от 02. 02.2021 № 5722-У «О формах, сроках и порядке составления и представления в Банк России отчетности и иных документов и информации кредитных потребительских кооперативов» (далее — Указание Банка России) и подлежащему отражению в графе 6 раздела XII в форме отчетности 0420820 «Отчет о деятельности кредитного потребительского кооператива» (далее — отчет о деятельности КПК) Указания Банка России).
02.2021 № 5722-У «О формах, сроках и порядке составления и представления в Банк России отчетности и иных документов и информации кредитных потребительских кооперативов» (далее — Указание Банка России) и подлежащему отражению в графе 6 раздела XII в форме отчетности 0420820 «Отчет о деятельности кредитного потребительского кооператива» (далее — отчет о деятельности КПК) Указания Банка России).
Согласно пункту 3 статьи 65.3 ГК РФ в случаях, в том числе предусмотренных другим законом, в корпорации образуется коллегиальный исполнительный орган (правление, дирекция и т.п.). К компетенции такого органа корпорации относится решение вопросов, не входящих в компетенцию ее высшего органа и коллегиального органа управления.
Так, в соответствии с положениями статьи 21 Закона № 190-ФЗ в периоды между общими собраниями членов КПК (пайщиков) руководство его деятельностью осуществляется правлением КПК. Члены правления КПК избираются общим собранием членов КПК (пайщиков) из числа членов КПК (пайщиков). При этом правление КПК не вправе решать вопросы, отнесенные Законом № 190-ФЗ и (или) уставом КПК к исключительной компетенции общего собрания членов КПК (пайщиков).
При этом правление КПК не вправе решать вопросы, отнесенные Законом № 190-ФЗ и (или) уставом КПК к исключительной компетенции общего собрания членов КПК (пайщиков).
С учетом изложенного правление является коллегиальным исполнительным органом КПК (информация о его членах отражается по коду основания аффилированности А1-2, установленному Указанием Банка России).
Также обращаем внимание, что в графе 6 раздела XII отчета о деятельности КПК Указания Банка России подлежит отражению информация о членах выборных органов КПК. Принимая во внимание, что согласно положениям части 3 статьи 24 Закона № 190-ФЗ персональный состав комитета по займам назначается правлением КПК, информация о членах комитета по займам не отражается в графе 6 раздела XII отчета о деятельности КПК.
женщин, сбитых грузовиком GK Morse на улице West 190, подали в суд на SDNY Discovery до 29 октября GK Morse Trucking на West 190 Street подает в суд Теперь SDNY Discovery до 29 октября
Мэтью
Рассел Ли, Патреон
BBC
— Страж
Великобритания — Гондурас
— ЕСПН
ЗДАНИЕ СУДА SDNY,
23 мая — Луизу Беато ранили. на автомобиле, принадлежащем GK Morse
Trucking Ltd 11 декабря,
2019 год на углу Запада
190-я улица и святой
Никольский проспект в Верхнем
Манхэттен.
на автомобиле, принадлежащем GK Morse
Trucking Ltd 11 декабря,
2019 год на углу Запада
190-я улица и святой
Никольский проспект в Верхнем
Манхэттен.
21 мая Окружной суд США
для Южного округа г.
Судья Нью-Йорка Эдгардо Рамос
провел судебное разбирательство. Внутренний город
Пресса освещала это.
Судья
Рамос сказал, что любые дополнительные
стороны должны присоединиться к июню
8, со всеми открытиями, сделанными
29 октября и дело
план управления в этот день. Есть спрос на жюри
пробный.
Есть спрос на жюри
пробный.
Чемодан Беато против GK Morse Trucking Ltd. и др. al., 20-cv-8291 (Рамос)
***
Ваш поддержка много значит.Всего 5 долларов в месяц
помогает нам продолжать работу и предоставляет вам доступ к
эксклюзивный бонусный материал на нашем Patreon
страница. Нажмите здесь, чтобы стать покровителем.
Обратная связь: Редакция [at] innercitypress.com
SDNY Press Room 480, передняя кабина
500 Pearl Street, NY NY 10007 USA
Почта: Box 20047, Dag
Станция Hammarskjold NY NY 10017
Мобильный корреспондент (и выходные): 718-716-3540
Другие, более ранние издания Inner City Press
перечислено здесь,
и некоторые из них доступны в ProQuest
сервис, а теперь и на Lexis-Nexis.
Авторское право 2006-2021 Внутренний город
Press, Inc. Для запроса перепечатки или других
разрешение, электронный контакт Редакция [at]
innercitypress.com
Коронарные проводники | Abbott
Коронарные проводники | ЭбботтОзнакомьтесь с постоянно растущим портфолио компании Abbott по направляющим тросам Workhorse и Specialty Wires.
Коронарные проводники Workhorse
Проволочный проводник, на который можно положиться при решении самых разных задач.Гибкий, с мягким наконечником и опорой для доставки большинства интервенционных устройств.
Специальные коронарные проводники
Решения для сложной анатомии и повреждений. Предлагаются различные нагрузки на наконечник, конусность наконечника, проникающая способность, покрытия и покрытия.
Предлагаются различные нагрузки на наконечник, конусность наконечника, проникающая способность, покрытия и покрытия.
ВЫ ХОТИТЕ ПРОДОЛЖИТЬ И ВЫХОДИТЬЭББОТ?
СОДЕРЖАНИЕ САЙТА НЕ НАХОДИТСЯ ПОД КОНТРОЛЕМ КОМПАНИИ ABBOTT.
Меры предосторожности
Обязательно прочитайте.
Следующие страницы предназначены для медицинских работников и содержат информацию о надлежащем использовании продуктов (медицинских устройств и т. д.) Abbott Medical Japan GK.
Представленная здесь информация не предназначена для предоставления информации пациентам и широкой общественности.
Вы медицинский работник?
Тест
да
Меры предосторожности
Обязательно прочитайте.
Следующие страницы предназначены для медицинских работников и содержат информацию о надлежащем использовании продуктов (медицинских устройств и т. д.) Abbott Medical Japan GK.
Представленная здесь информация не предназначена для предоставления информации пациентам и широкой общественности.
Вы медицинский работник?
Тест
[продукт, crx3, образец содержимого, публикация, crx3tar]
ВЫ ХОТИТЕ ПРОДОЛЖИТЬ И ВЫХОДИТЬЭББОТ?
СОДЕРЖАНИЕ САЙТА НЕ НАХОДИТСЯ ПОД КОНТРОЛЕМ КОМПАНИИ ABBOTT.
https://сосудистый. abbott.com/,https://mri.merlin.net/,https://www.thelancet.com/,https://www.ahajournals.org/,https://www.onlinejacc.org/,https ://jamanetwork.com/,https://www.sciencedirect.com/,https://onlinelibrary.wiley.com/,https://www.cms.gov/,https://www.novitas-solutions .com/,https://event.on24.com/,https://dx.doi.org/,https://www.myloopaccount.com/,https://www.invivecardiology.com/,https: //manuals.sjm.com/,https://www.кардиоваскулярный.abbott
abbott.com/,https://mri.merlin.net/,https://www.thelancet.com/,https://www.ahajournals.org/,https://www.onlinejacc.org/,https ://jamanetwork.com/,https://www.sciencedirect.com/,https://onlinelibrary.wiley.com/,https://www.cms.gov/,https://www.novitas-solutions .com/,https://event.on24.com/,https://dx.doi.org/,https://www.myloopaccount.com/,https://www.invivecardiology.com/,https: //manuals.sjm.com/,https://www.кардиоваскулярный.abbott
Ложь
доступность© 2021 Эбботт.Все права защищены. Пожалуйста, ознакомьтесь с официальным уведомлением для получения дополнительной информации.
Если не указано иное, все названия продуктов и услуг, встречающиеся на этом интернет-сайте, являются товарными знаками, принадлежащими или лицензированными компанией Abbott, ее дочерними или аффилированными компаниями. Никакое использование товарных знаков, торговых наименований или фирменного стиля Abbott на этом сайте не допускается без предварительного письменного разрешения Abbott, кроме как для идентификации продукции или услуг компании.
ВЫ ХОТИТЕ ПРОДОЛЖИТЬ И ВЫХОДИТЬЭББОТ?
СОДЕРЖАНИЕ САЙТА НЕ НАХОДИТСЯ ПОД КОНТРОЛЕМ КОМПАНИИ ABBOTT.
доступность© 2021 Эбботт.Все права защищены. Пожалуйста, ознакомьтесь с официальным уведомлением для получения дополнительной информации.
Если не указано иное, все названия продуктов и услуг, встречающиеся на этом интернет-сайте, являются товарными знаками, принадлежащими или лицензированными компанией Abbott, ее дочерними или аффилированными компаниями. Никакое использование товарных знаков, торговых наименований или фирменного стиля Abbott на этом сайте не допускается без предварительного письменного разрешения Abbott, кроме как для идентификации продукции или услуг компании.
доступностьМеры предосторожности
Обязательно прочитайте.
Следующие страницы предназначены для медицинских работников и содержат информацию о надлежащем использовании продуктов (медицинских устройств и т. д.) Abbott Medical Japan GK.
д.) Abbott Medical Japan GK.
Представленная здесь информация не предназначена для предоставления информации пациентам и широкой общественности.
Вы медицинский работник?
Тест
Этот сайт использует файлы cookie.Используя этот сайт, вы даете согласие на размещение наших файлов cookie. Прочтите нашу политику конфиденциальности, чтобы узнать больше.
границ | Мышечная стволовая клетка для каждой мышцы: вариабельность биологии сателлитных клеток в разных группах мышц
Введение
Скелетные мышцы представляют собой высокоорганизованную ткань, которая составляет до 40% массы тела человека и необходима для основных функций, таких как обмен веществ, передвижение и дыхание (Janssen et al. , 2000; Макинтош и др., 2006). Человеческое тело содержит около 640 уникальных скелетных мышц, каждая из которых выполняет определенные функции в физиологии человека. Многочисленные факторы способствуют разнообразию скелетных мышц, включая эмбриологическое происхождение, пути миогенной регуляции и функциональные/метаболические потребности. Мышечная гетерогенность дополнительно подчеркивается различной чувствительностью конкретных подмножеств скелетных мышц к многочисленным различным генетическим мутациям, которые вызывают мышечные дистрофии (MD; Hoffman et al., 1987; Монако и др., 1988; Бионе и др., 1994; Брайс и др., 1998; Бонн и др., 1999; Нонака, 1999 г.; Эмери, 2002 г.; Робинсон и др., 2005 г.; Боннеманн и др., 2014; Виейра и др., 2014). Взрослые мышечные стволовые клетки, называемые сателлитными клетками, вовлечены в патологию некоторых МД (таблица 1) и могут способствовать вариабельной мышечной чувствительности, наблюдаемой при некоторых дистрофиях.
, 2000; Макинтош и др., 2006). Человеческое тело содержит около 640 уникальных скелетных мышц, каждая из которых выполняет определенные функции в физиологии человека. Многочисленные факторы способствуют разнообразию скелетных мышц, включая эмбриологическое происхождение, пути миогенной регуляции и функциональные/метаболические потребности. Мышечная гетерогенность дополнительно подчеркивается различной чувствительностью конкретных подмножеств скелетных мышц к многочисленным различным генетическим мутациям, которые вызывают мышечные дистрофии (MD; Hoffman et al., 1987; Монако и др., 1988; Бионе и др., 1994; Брайс и др., 1998; Бонн и др., 1999; Нонака, 1999 г.; Эмери, 2002 г.; Робинсон и др., 2005 г.; Боннеманн и др., 2014; Виейра и др., 2014). Взрослые мышечные стволовые клетки, называемые сателлитными клетками, вовлечены в патологию некоторых МД (таблица 1) и могут способствовать вариабельной мышечной чувствительности, наблюдаемой при некоторых дистрофиях.
ТАБЛИЦА 1. Мышечные дистрофии (МД), при которых дефицит функции сателлитных клеток связан с патологией заболевания.
Мышечные дистрофии (МД), при которых дефицит функции сателлитных клеток связан с патологией заболевания.
Скелетные мышцы состоят из миофибрилл, крупных синцитиальных клеток, содержащих сотни постмитотических миоядер. Расположенные между базальной пластинкой и клеточной мембраной миофибрилл, сателлитные клетки располагаются на периферии скелетных миофибрилл (Mauro, 1961). Недавние исследования показали, что сателлитные клетки, экспрессирующие парный белок 7 (Pax7) , являются первичными миогенными клетками, необходимыми для регенерации мышц (Lepper et al., 2011; Murphy et al., 2011; Sambasivan et al., 2011). Большая часть знаний о биологии сателлитных клеток получена в результате изучения мышц, происходящих из сомитов, таких как четырехглавая, икроножная, передняя большеберцовая (TA), камбаловидная мышца, длинный разгибатель пальцев, подошвенная, двуглавая и дельтовидная мышцы, которые в совокупности составляют менее 2% всех скелетных мышц. Интересно, что сателлитные клетки, присутствующие в других группах мышц, включая туловище, диафрагму, гортань, язык, экстраокулярные, жевательные мышцы и глотку, отличаются от канонической биологии своих аналогов конечностей. Здесь мы обращаемся к мышечно-специфической изменчивости биологии сателлитных клеток и постулируем, как эта изменчивость может способствовать мышечно-специфической чувствительности, обнаруживаемой при MD.
Здесь мы обращаемся к мышечно-специфической изменчивости биологии сателлитных клеток и постулируем, как эта изменчивость может способствовать мышечно-специфической чувствительности, обнаруживаемой при MD.
Сателлитные клетки мышц конечностей: создание Canon
Скелетные мышцы задних конечностей грызунов обычно используются для изучения сателлитных клеток, поскольку эти мышцы легко идентифицировать, препарировать, собирать и манипулировать экспериментально. Скелетные мышцы конечностей и живота возникают из сомитовой мезодермы и называются гипаксиальными мышцами (рис. 1).Они возникают в процессе развития из вентролатерального дермомиотома сегментированной параксиальной мезодермы. Исследования in vivo и in vitro , изучающие мышцы конечностей, дают фундаментальное представление о механизмах и регуляторных путях, связанных с регенерацией скелетных мышц, ростом мышц и биологией сателлитных клеток.
РИСУНОК 1. Эмбриональные мезодермальные вклады в скелетные мышцы взрослых. (A) Схема мезодермального происхождения у эмбриона мыши на стадии 3–5 сомитов. (B) Скелетные мышцы туловища, конечностей, диафрагмы и языка возникают из сомитной мезодермы. Напротив, экстраокулярные мышцы (EOM) возникают из прехордальной мезодермы и краниальной параксиальной мезодермы первой глоточной дуги; жевательная мышца от первой и второй глоточных дуг краниальной параксиальной мезодермы и глотка от третьей и четвертой глоточных дуг каудальной параксиальной мезодермы. Мышцы языка возникают как из сомитной, так и из краниальной мезодермы, развиваясь в нише краниальной мезенхимы, которая снабжается всеми четырьмя глоточными дугами.
(A) Схема мезодермального происхождения у эмбриона мыши на стадии 3–5 сомитов. (B) Скелетные мышцы туловища, конечностей, диафрагмы и языка возникают из сомитной мезодермы. Напротив, экстраокулярные мышцы (EOM) возникают из прехордальной мезодермы и краниальной параксиальной мезодермы первой глоточной дуги; жевательная мышца от первой и второй глоточных дуг краниальной параксиальной мезодермы и глотка от третьей и четвертой глоточных дуг каудальной параксиальной мезодермы. Мышцы языка возникают как из сомитной, так и из краниальной мезодермы, развиваясь в нише краниальной мезенхимы, которая снабжается всеми четырьмя глоточными дугами.
Регенерация мышц — это надежный и сложный клеточный процесс, который восстанавливает поврежденную мышцу до состояния, морфологически и функционально аналогичного неповрежденной мышце (рис. 2; Abmayr and Pavlath, 2012). Регенерация скелетных мышц происходит в две отдельные фазы: дегенеративную фазу и регенеративную фазу (Rai et al. , 2014). Основные характеристики дегенеративной фазы включают повреждение сарколеммы миофибрилл или некроз миофибрилл с последующим притоком мононуклеарных воспалительных клеток и увеличением числа фибробластов (Mathew et al., 2011; Мерфи и др., 2011 г.; Рай и др., 2014). Факторы, высвобождаемые из поврежденных мышечных волокон, инициируют воспалительную реакцию, которая привлекает нейтрофилы, макрофаги и активирует фиброзные/адипогенные предшественники, чтобы облегчить удаление клеточного дебриса и регулировать восстановление мышц (McLennan, 1996; Lescaudron et al., 1999; Joe et al., 2010). ; Уезуми и др., 2010; Паллафаччина и др., 2013). Базальная пластинка остается интактной, выступая в качестве каркаса для следующей фазы регенерации мышц (Schmalbruch, 1976). Высвобождаются несколько молекулярных сигналов, таких как факторы роста, хемокины и цитокины, которые активируют сателлитные клетки как локально, так и системно в течение первых 24–48 часов после травмы (Chang and Rudnicki, 2014; Rodgers et al.
, 2014). Основные характеристики дегенеративной фазы включают повреждение сарколеммы миофибрилл или некроз миофибрилл с последующим притоком мононуклеарных воспалительных клеток и увеличением числа фибробластов (Mathew et al., 2011; Мерфи и др., 2011 г.; Рай и др., 2014). Факторы, высвобождаемые из поврежденных мышечных волокон, инициируют воспалительную реакцию, которая привлекает нейтрофилы, макрофаги и активирует фиброзные/адипогенные предшественники, чтобы облегчить удаление клеточного дебриса и регулировать восстановление мышц (McLennan, 1996; Lescaudron et al., 1999; Joe et al., 2010). ; Уезуми и др., 2010; Паллафаччина и др., 2013). Базальная пластинка остается интактной, выступая в качестве каркаса для следующей фазы регенерации мышц (Schmalbruch, 1976). Высвобождаются несколько молекулярных сигналов, таких как факторы роста, хемокины и цитокины, которые активируют сателлитные клетки как локально, так и системно в течение первых 24–48 часов после травмы (Chang and Rudnicki, 2014; Rodgers et al. , 2014). Затем миобласты окончательно дифференцируются, превращаясь в постмитотические миоциты, которые затем сливаются с другими миоцитами или миофибриллами для регенерации или восстановления поврежденных миофибрилл. Таким образом, новые миоядра добавляются к поврежденным или зарождающимся миофибриллам (Abmayr and Pavlath, 2012). Субпопуляция миогенных клеток повторно заселяет нишу сателлитных клеток, тем самым поддерживая и пополняя пул покоящихся сателлитных клеток для последующих циклов регенерации (Collins et al., 2005; Shinin et al., 2006).
, 2014). Затем миобласты окончательно дифференцируются, превращаясь в постмитотические миоциты, которые затем сливаются с другими миоцитами или миофибриллами для регенерации или восстановления поврежденных миофибрилл. Таким образом, новые миоядра добавляются к поврежденным или зарождающимся миофибриллам (Abmayr and Pavlath, 2012). Субпопуляция миогенных клеток повторно заселяет нишу сателлитных клеток, тем самым поддерживая и пополняя пул покоящихся сателлитных клеток для последующих циклов регенерации (Collins et al., 2005; Shinin et al., 2006).
РИСУНОК 2. Структура миофибрилл и клеточная прогрессия миогенеза. Миофибриллы окружены базальной пластинкой, под которой лежат сателлитные клетки, плотно прилегающие к миофибриллам. При повреждении сателлитные клетки пролиферируют и дают начало миобластам, которые дифференцируются, мигрируют, прилипают и сливаются друг с другом, образуя множественные мышечные трубки в каркасе базальной пластинки. Миобласты/миотрубочки сливаются с культями выживших миофибрилл, и миотрубочки также сливаются друг с другом для восстановления поврежденных миофибрилл. Регенерированные миофибриллы идентифицируются по наличию центрально расположенных ядер. Репрезентативные окрашенные гематоксилином и эозином поперечные срезы мышц химически поврежденных мышц мышей представлены для каждой стадии регенерации мышц, чтобы проиллюстрировать дифференциальную морфологию тканей.
Регенерированные миофибриллы идентифицируются по наличию центрально расположенных ядер. Репрезентативные окрашенные гематоксилином и эозином поперечные срезы мышц химически поврежденных мышц мышей представлены для каждой стадии регенерации мышц, чтобы проиллюстрировать дифференциальную морфологию тканей.
Роль сателлитных клеток в постнатальном росте изучалась также в мышцах конечностей. У мышей первые 3 недели неонатального роста приводят к трехкратному увеличению мышечной массы, в течение которого популяция сателлитных клеток подвергается значительному сокращению с ~ 30% миоядер на миоволокно до 5% после слияния с неонатальными мышцами.Параллельное увеличение числа миоядер и цитоплазматических белков происходит до 21 дня постнатального развития (White et al., 2010). После постнатального дня 21 сателлитные клетки переходят в состояние покоя под регуляцией передачи сигналов Notch (Fukada et al., 2011), но размер миофибрилл продолжает увеличиваться без добавления новых миоядер (White et al. , 2010). Недавние исследования по удалению сателлитных клеток также показали, что миоядерное добавление из сателлитных клеток необязательно для гипертрофического роста мышц конечностей у взрослых (McCarthy et al., 2011). Кроме того, сателлитные клетки, по-видимому, не требуются для поддержания большинства мышц конечностей взрослого человека. Недавнее исследование абляции сателлитных клеток показало, что потеря >90% сателлитных клеток взрослых конечностей не приводит к изменению размера мышц или типа миофибрилл в пяти различных мышцах конечностей с возрастом (Fry et al., 2015). Тем не менее, миоядерное добавление происходит на базальном уровне в неповрежденных постнатальных мышцах конечностей и может быть необходимо для поддержания размера миофибрилл длинного разгибателя пальцев с возрастом (Keefe et al., 2015). В совокупности эти исследования предполагают, что начальная фаза постнатального роста мышц происходит с добавлением миоядер из сателлитных клеток, но поддержание размера мышц большинства взрослых конечностей не зависит от сателлитных клеток.
, 2010). Недавние исследования по удалению сателлитных клеток также показали, что миоядерное добавление из сателлитных клеток необязательно для гипертрофического роста мышц конечностей у взрослых (McCarthy et al., 2011). Кроме того, сателлитные клетки, по-видимому, не требуются для поддержания большинства мышц конечностей взрослого человека. Недавнее исследование абляции сателлитных клеток показало, что потеря >90% сателлитных клеток взрослых конечностей не приводит к изменению размера мышц или типа миофибрилл в пяти различных мышцах конечностей с возрастом (Fry et al., 2015). Тем не менее, миоядерное добавление происходит на базальном уровне в неповрежденных постнатальных мышцах конечностей и может быть необходимо для поддержания размера миофибрилл длинного разгибателя пальцев с возрастом (Keefe et al., 2015). В совокупности эти исследования предполагают, что начальная фаза постнатального роста мышц происходит с добавлением миоядер из сателлитных клеток, но поддержание размера мышц большинства взрослых конечностей не зависит от сателлитных клеток.
Регуляторные гены, участвующие в биологии сателлитных клеток, также были выявлены в результате исследований мышц конечностей. Во взрослых скелетных мышцах покоящиеся сателлитные клетки экспрессируют Pax7, фактор транскрипции, который специфицирует миогенное происхождение (Seale et al., 2000).После активации сателлитные клетки выходят из клеточного покоя, вступают в клеточный цикл и начинают прогрессировать по миогенной линии под контролем миогенных регуляторных факторов (MRF), специфических для мышц факторов транскрипции класса основная спираль-петля-спираль (bHLH). , включая белок миогенной дифференцировки (MyoD), миогенный фактор 5 (Myf5), миогенный регуляторный фактор 4 (Mrf4) и миогенин (Weintraub et al., 1991; Olson and Klein, 1994; Chang and Rudnicki, 2014). MyoD и Myf5 экспрессируются во время пролиферативной фазы и регулируют миогенную дифференцировку (Cooper et al., 1999; Valdez et al., 2000), тогда как Mrf4 и миогенин экспрессируются при терминальной дифференцировке и выходе из клеточного цикла (Chang, Rudnicki, 2014).
Все больше данных свидетельствует о том, что сателлитные клетки в мышцах гетерогенны (Motohashi and Asakura, 2014). Сателлитные клетки, содержащие высокий уровень Pax7, демонстрируют более медленную скорость пролиферации, более низкий метаболизм и устойчивость к дифференцировке, что указывает на более «стволовой» фенотип по сравнению с сателлитными клетками с более низким уровнем Pax7 (Rocheteau et al., 2012). Различные группы также обнаружили различные субпопуляции сателлитных клеток на основе дифференциальной экспрессии других белков, включая α7-интегрин, β1-интегрин, c-met, CD34, рецептор кальцитонина, хемокиновый рецептор CXC типа 4 (CXCR4), M-кадгерин, Myf5, нейральный молекула клеточной адгезии 1, синдеканы 3 и 4 и молекула сосудистой клеточной адгезии 1 (Rosen et al., 1992; Cornelison and Wold, 1997; Beauchamp et al., 2000; Blanco-Bose et al., 2001; Cornelison et al. , 2001; Тамаки и др., 2002; Шервуд и др., 2004; Фукада и др., 2007 г.; Икемото и др., 2007 г.; Куанг и др. , 2007 г.; Кафадар и др., 2009). В то время как механизмы, лежащие в основе гетерогенности сателлитных клеток, все еще выясняются, все больше данных свидетельствует о том, что биология сателлитных клеток также зависит от мышц, как обсуждается ниже.
, 2007 г.; Кафадар и др., 2009). В то время как механизмы, лежащие в основе гетерогенности сателлитных клеток, все еще выясняются, все больше данных свидетельствует о том, что биология сателлитных клеток также зависит от мышц, как обсуждается ниже.
«Другие» мышцы, происходящие из сомитов: эпиаксиальные, диафрагмальные, внутренние клетки гортани и сателлитные клетки языка
Эпаксиальные сателлитные ячейки
Эпаксиальные скелетные мышцы включают глубокие мышцы спины.Подобно развитию гипаксиальных мышц, эпаксиальные мышцы возникают из дорсомедиального дермомиотома сегментированной параксиальной мезодермы (Рис. 1; Christ and Ordahl, 1995; Burke and Nowicki, 2003). В то время как развитие эпаксиальной мышцы хорошо изучено (Munsterberg et al., 1995; Tajbakhsh et al., 1997; Borycki et al., 1999; Gustafsson et al., 2002; McDermott et al., 2005; Borello et al., 2006; L’Honore et al., 2010; Sato et al., 2010), биологические свойства взрослых эпаксиальных сателлитных клеток остаются в значительной степени неизвестными. Модели мышей, экспрессирующих nLacZ под контролем промотора Pax3 и , показали, что коэкспрессия Pax3 и Pax7 сохраняется в большинстве взрослых сателлитных клеток глубоких вентральных мышц туловища (Relaix et al., 2006). Экспрессия Pax3 также поддерживалась в культивируемых эпаксиальных миобластах, подтверждая, что Pax3 может вносить вклад как в состояние покоя, так и в активированное состояние эпаксиальных сателлитных клеток (Relaix et al., 2006). Как Pax3 вносит вклад в биологию эпаксиальных сателлитных клеток взрослых, и роль Pax3 в регенерации эпаксиальных мышц еще предстоит определить, так же как и др. фундаментальные аспекты биологии эваксиальных сателлитных клеток и миогенеза.
Модели мышей, экспрессирующих nLacZ под контролем промотора Pax3 и , показали, что коэкспрессия Pax3 и Pax7 сохраняется в большинстве взрослых сателлитных клеток глубоких вентральных мышц туловища (Relaix et al., 2006). Экспрессия Pax3 также поддерживалась в культивируемых эпаксиальных миобластах, подтверждая, что Pax3 может вносить вклад как в состояние покоя, так и в активированное состояние эпаксиальных сателлитных клеток (Relaix et al., 2006). Как Pax3 вносит вклад в биологию эпаксиальных сателлитных клеток взрослых, и роль Pax3 в регенерации эпаксиальных мышц еще предстоит определить, так же как и др. фундаментальные аспекты биологии эваксиальных сателлитных клеток и миогенеза.
Мембранные сателлитные ячейки
Мышца диафрагмы состоит из трех отдельных доменов: мышцы голени, реберной мышцы и центрального сухожильного домена (Anraku and Shargall, 2009). Недавние исследования свидетельствуют о том, что мышечные компоненты возникают из латерального дермомиотома шейных сомитов, тогда как центральная сухожильная соединительная ткань возникает из плеевроперитонеальных складок латеральной пластинки (рис. 1; Noden et al., 1999; Mootoosamy and Dietrich, 2002; Pickering). and Jones, 2002; Бабюк и др., 2003; Браун и др., 2005 г.; Меррелл и др., 2015). Постнатально диафрагмальные сателлитные клетки отличаются от гипаксиальных мышц тем, что экспрессия Pax3 сохраняется наряду с Pax7 и Myf5 (Relaix et al., 2006; Day et al., 2007; Stuelsatz et al., 2012). Недавние исследования показывают, что происходящие из сателлитных клеток миобласты диафрагмы ведут себя иначе, чем миобласты задних конечностей in vitro . Используя модель мыши Nestin-GFP (Day et al., 2007) для идентификации самообновления миобластов, Stuelsatz et al.(2014) обнаружил более высокий процент клеток GFP + в мышечных культурах в диафрагме. Анализы клональной экспансии in vitro показали, что диафрагмальные миобласты пролиферируют так же, как и мышцы задних конечностей (Chen et al., 2011; Stuelsatz et al., 2012), но миогенная дифференцировка культур диафрагмы постоянно достигала максимальных показателей слияния раньше, чем культуры конечностей (Chen et al.
1; Noden et al., 1999; Mootoosamy and Dietrich, 2002; Pickering). and Jones, 2002; Бабюк и др., 2003; Браун и др., 2005 г.; Меррелл и др., 2015). Постнатально диафрагмальные сателлитные клетки отличаются от гипаксиальных мышц тем, что экспрессия Pax3 сохраняется наряду с Pax7 и Myf5 (Relaix et al., 2006; Day et al., 2007; Stuelsatz et al., 2012). Недавние исследования показывают, что происходящие из сателлитных клеток миобласты диафрагмы ведут себя иначе, чем миобласты задних конечностей in vitro . Используя модель мыши Nestin-GFP (Day et al., 2007) для идентификации самообновления миобластов, Stuelsatz et al.(2014) обнаружил более высокий процент клеток GFP + в мышечных культурах в диафрагме. Анализы клональной экспансии in vitro показали, что диафрагмальные миобласты пролиферируют так же, как и мышцы задних конечностей (Chen et al., 2011; Stuelsatz et al., 2012), но миогенная дифференцировка культур диафрагмы постоянно достигала максимальных показателей слияния раньше, чем культуры конечностей (Chen et al. , 2011; Stuelsatz et al., 2012) Stuelsatz и др., 2014). Однако другие исследования показали, что сателлитные клетки диафрагмы подвергаются повышенной пролиферации и пониженной дифференцировке in vitro по сравнению с мышцами конечностей, туловища и черепно-лицевых мышц (Ippolito et al., 2012). Расхождения в этих исследованиях свидетельствуют о необходимости дальнейшего изучения биологии сателлитных клеток диафрагмы. Исследования in vivo также указывают на некоторую интересную биологию, связанную с диафрагмальными сателлитными клетками. In vivo исследований по отслеживанию клонов, изучающих вклад диафрагмальных сателлитных клеток в неповрежденные диафрагмальные мышцы, выявили продолжающееся добавление новых миоядер с возрастом, однако абляция сателлитных клеток не влияла на размер миофибрилл (Keefe et al., 2015). Одно исследование на крысах предполагает, что существует гетерогенность в ответе клеток-сателлитов диафрагмы на стимулирующие рост стимулы, обеспечиваемые денервацией полудиафрагмы.
, 2011; Stuelsatz et al., 2012) Stuelsatz и др., 2014). Однако другие исследования показали, что сателлитные клетки диафрагмы подвергаются повышенной пролиферации и пониженной дифференцировке in vitro по сравнению с мышцами конечностей, туловища и черепно-лицевых мышц (Ippolito et al., 2012). Расхождения в этих исследованиях свидетельствуют о необходимости дальнейшего изучения биологии сателлитных клеток диафрагмы. Исследования in vivo также указывают на некоторую интересную биологию, связанную с диафрагмальными сателлитными клетками. In vivo исследований по отслеживанию клонов, изучающих вклад диафрагмальных сателлитных клеток в неповрежденные диафрагмальные мышцы, выявили продолжающееся добавление новых миоядер с возрастом, однако абляция сателлитных клеток не влияла на размер миофибрилл (Keefe et al., 2015). Одно исследование на крысах предполагает, что существует гетерогенность в ответе клеток-сателлитов диафрагмы на стимулирующие рост стимулы, обеспечиваемые денервацией полудиафрагмы. Изучая количество сателлитных клеток, связанных с различными типами волокон, это исследование показало, что только миофибриллы типа IIx/b компенсирующей неденервированной гемидиафрагмы показали значительное увеличение количества сателлитных клеток как у молодых, так и у взрослых мышей, в то время как никаких изменений в количестве не было. связаны с миофибриллами типа I или IIa (Kawai et al., 2012). В совокупности наше текущее понимание биологии сателлитных клеток диафрагмы все еще находится в зачаточном состоянии и требует дальнейшего изучения, как in vivo , так и in vitro .
Изучая количество сателлитных клеток, связанных с различными типами волокон, это исследование показало, что только миофибриллы типа IIx/b компенсирующей неденервированной гемидиафрагмы показали значительное увеличение количества сателлитных клеток как у молодых, так и у взрослых мышей, в то время как никаких изменений в количестве не было. связаны с миофибриллами типа I или IIa (Kawai et al., 2012). В совокупности наше текущее понимание биологии сателлитных клеток диафрагмы все еще находится в зачаточном состоянии и требует дальнейшего изучения, как in vivo , так и in vitro .
Внутренние сателлитные клетки гортани
Собственные мышцы гортани имеют решающее значение для дыхания, защиты дыхательных путей и вокализации и включают щиточерпаловидную, заднюю перстнечерпаловидную (PCA) и латеральную перстнечерпаловидную мышцы. Собственные мышцы гортани возникают из затылочных сомитов во время развития (рис. 1; Noden et al., 1999; Ноден и Фрэнсис-Уэст, 2006). В тиреочерпаловидных мышцах и мышцах PCA базальные уровни миоядерного добавления были обнаружены с использованием отслеживания клонов BrdU, подтверждая, что неповрежденная гортанная мышца содержит субнабор пролиферирующих сателлитных клеток (Goding et al., 2005). Исследования in vitro также обнаружили повышенную скорость пролиферации, связанную с фосфорилированием Erk1/2, в первичных культурах мышц гортани по сравнению с культурами задних конечностей (Walz et al., 2008). В ответ на денервацию гортани происходит активация и пролиферация сателлитных клеток гортани in vivo в течение нескольких дней после денервации (Kumai et al., 2007) с добавлением миоядер, происходящим во всех проанализированных внутренних мышцах гортани (Shinners et al., 2006). Удивительно, но миогенные клетки, экспрессирующие MyoD и myogenin, все еще присутствуют в тиреочерпаловидных мышцах через 2 года после денервации у людей, что свидетельствует об уникальном пролонгированном регенеративном потенциале в мышцах гортани (Donghui et al.
В тиреочерпаловидных мышцах и мышцах PCA базальные уровни миоядерного добавления были обнаружены с использованием отслеживания клонов BrdU, подтверждая, что неповрежденная гортанная мышца содержит субнабор пролиферирующих сателлитных клеток (Goding et al., 2005). Исследования in vitro также обнаружили повышенную скорость пролиферации, связанную с фосфорилированием Erk1/2, в первичных культурах мышц гортани по сравнению с культурами задних конечностей (Walz et al., 2008). В ответ на денервацию гортани происходит активация и пролиферация сателлитных клеток гортани in vivo в течение нескольких дней после денервации (Kumai et al., 2007) с добавлением миоядер, происходящим во всех проанализированных внутренних мышцах гортани (Shinners et al., 2006). Удивительно, но миогенные клетки, экспрессирующие MyoD и myogenin, все еще присутствуют в тиреочерпаловидных мышцах через 2 года после денервации у людей, что свидетельствует об уникальном пролонгированном регенеративном потенциале в мышцах гортани (Donghui et al. , 2010). Несмотря на это, происходит возрастное снижение плотности сателлитных клеток гортани и нарушение регенерации щиточерпаловидных мышц (Malmgren et al., 2000; Lee et al., 2012). Вместе эти исследования обеспечивают интригующую информацию о внутренних сателлитных клетках гортани, но необходимы дальнейшие исследования для выяснения молекулярных и регуляторных механизмов, лежащих в основе их особой биологии.
, 2010). Несмотря на это, происходит возрастное снижение плотности сателлитных клеток гортани и нарушение регенерации щиточерпаловидных мышц (Malmgren et al., 2000; Lee et al., 2012). Вместе эти исследования обеспечивают интригующую информацию о внутренних сателлитных клетках гортани, но необходимы дальнейшие исследования для выяснения молекулярных и регуляторных механизмов, лежащих в основе их особой биологии.
Сателлитные клетки языка
Мышцы языка возникают из мезодермального происхождения смешанного происхождения. Собственные мышцы возникают из первого затылочного сомита, а параксиальная мезодерма способствует формированию наружных мышц языка (рис. 1; Shuler and Dalrymple, 2001; Czajkowski et al., 2014). На сегодняшний день знания о биологии сателлитных клеток мышц языка взрослых сильно ограничены. Исследование денервации мышцы языка с использованием 3 H-тимидина для мечения пролиферирующих клеток выявило увеличение количества миоядер, меченных 3 H-тимидином, что свидетельствует о слиянии пролиферирующих сателлитных клеток с миофибриллами (McGeachie and Allbrook, 1978). Миобласты, происходящие из языка, были выделены, культивированы и успешно дифференцированы в зарождающиеся миотрубочки 90–211 in vitro 90–212, но миогенные характеристики этих клеток еще предстоит изучить напрямую (Ternaux and Portalier, 1993; LaFramboise et al., 2003).
Миобласты, происходящие из языка, были выделены, культивированы и успешно дифференцированы в зарождающиеся миотрубочки 90–211 in vitro 90–212, но миогенные характеристики этих клеток еще предстоит изучить напрямую (Ternaux and Portalier, 1993; LaFramboise et al., 2003).
Черепно-лицевые сателлитные клетки: экстраокулярные, жевательные и глоточные
Экстраокулярные сателлитные клетки
Экстраокулярные мышцы (ЭГМ) отвечают за вращение и движение глаза и включают верхнюю косую, нижнюю косую, верхнюю прямую, нижнюю прямую, латеральную прямую и медиальную прямую мышцы. ЭОМ и связанные с ними сателлитные клетки отличаются от других скелетных мышц тем, что они возникают из прехордальной и краниальной параксиальной мезодермы первой глоточной дуги во время развития (рис. 1; Couly et al., 1992; Ноден и Фрэнсис-Уэст, 2006). Ранние исследования, посвященные изучению последствий старения и дистрофических заболеваний, показали, что мышцы EOM отличаются от своих аналогов задних конечностей. Исследования старения показали, что ЭСО преимущественно сохраняются с возрастом (Porter et al., 1998; Schoser and Pongratz, 2006), в то время как исследования, изучающие участие ЭСО при МД Дюшенна, Беккера и некоторых заболеваниях пояса конечностей, также показали преимущественное сохранение этих мышц (Kaminski и др., 1992; Эмери, 2002). Кроме того, сателлитные клетки EOM имеют уникальные профили экспрессии генов по сравнению с покоящимися сателлитными клетками мышц задних конечностей (Porter and Baker, 1996; Pacheco-Pinedo et al., 2009). Сателлитные клетки EOM также демонстрируют отчетливые биологические различия по сравнению с сателлитными клетками задних конечностей. У некоторых видов сателлитные клетки EOM хронически пролиферируют in vivo (McLoon and Wirtschafter, 2002a, 2003; Wirtschafter et al., 2004; Christiansen and McLoon, 2006), что отчасти может быть связано со специфической высокопролиферативной субпопуляцией (Kallestad). и др., 2011). Фактор транскрипции Pitx2 экспрессируется в постнатальных миогенных клетках-предшественниках ЭОМ, принадлежащих к линии CD34 + /Sca1 — /CD31 — /CD45 — , что, как считается, способствует пролиферативным свойствам ЭОМ.
Исследования старения показали, что ЭСО преимущественно сохраняются с возрастом (Porter et al., 1998; Schoser and Pongratz, 2006), в то время как исследования, изучающие участие ЭСО при МД Дюшенна, Беккера и некоторых заболеваниях пояса конечностей, также показали преимущественное сохранение этих мышц (Kaminski и др., 1992; Эмери, 2002). Кроме того, сателлитные клетки EOM имеют уникальные профили экспрессии генов по сравнению с покоящимися сателлитными клетками мышц задних конечностей (Porter and Baker, 1996; Pacheco-Pinedo et al., 2009). Сателлитные клетки EOM также демонстрируют отчетливые биологические различия по сравнению с сателлитными клетками задних конечностей. У некоторых видов сателлитные клетки EOM хронически пролиферируют in vivo (McLoon and Wirtschafter, 2002a, 2003; Wirtschafter et al., 2004; Christiansen and McLoon, 2006), что отчасти может быть связано со специфической высокопролиферативной субпопуляцией (Kallestad). и др., 2011). Фактор транскрипции Pitx2 экспрессируется в постнатальных миогенных клетках-предшественниках ЭОМ, принадлежащих к линии CD34 + /Sca1 — /CD31 — /CD45 — , что, как считается, способствует пролиферативным свойствам ЭОМ. сателлитные клетки (Hebert et al., 2013). Кроме того, состарившиеся сателлитные клетки EOM сохраняют способность к пролиферации и самообновлению вплоть до 24-месячного возраста in vitro (Stuelsatz et al., 2014). Кроме того, глобальные и орбитальные сателлитные клетки EOM вносят новые миоядра в миофибриллы EOM при отсутствии повреждений (McLoon and Wirtschafter, 2002a,b, 2003; Wirtschafter et al., 2004; Keefe et al., 2015). Вместе эти исследования подчеркивают биологию сателлитных клеток, которая отличается от сателлитных клеток задних конечностей. Из-за их склонности к пролиферации и самообновлению сателлитные клетки EOM были предложены в качестве идеальных кандидатов для использования в клеточной терапии миопатического заболевания (McLoon et al., 2007; Каллестад и др., 2011; Stuelsatz и др., 2014). Однако в экспериментах по трансплантации, в которых сателлитные клетки EOM вводили в TA мышцу задней конечности, сателлитные клетки EOM успешно приживались в новой нише, но теряли специфические для EOM фенотипы, такие как экспрессия Myh23 и медленно тонического миозина (Sambasivan et al.
сателлитные клетки (Hebert et al., 2013). Кроме того, состарившиеся сателлитные клетки EOM сохраняют способность к пролиферации и самообновлению вплоть до 24-месячного возраста in vitro (Stuelsatz et al., 2014). Кроме того, глобальные и орбитальные сателлитные клетки EOM вносят новые миоядра в миофибриллы EOM при отсутствии повреждений (McLoon and Wirtschafter, 2002a,b, 2003; Wirtschafter et al., 2004; Keefe et al., 2015). Вместе эти исследования подчеркивают биологию сателлитных клеток, которая отличается от сателлитных клеток задних конечностей. Из-за их склонности к пролиферации и самообновлению сателлитные клетки EOM были предложены в качестве идеальных кандидатов для использования в клеточной терапии миопатического заболевания (McLoon et al., 2007; Каллестад и др., 2011; Stuelsatz и др., 2014). Однако в экспериментах по трансплантации, в которых сателлитные клетки EOM вводили в TA мышцу задней конечности, сателлитные клетки EOM успешно приживались в новой нише, но теряли специфические для EOM фенотипы, такие как экспрессия Myh23 и медленно тонического миозина (Sambasivan et al. ., 2009). Эти данные свидетельствуют о том, что фенотипы сателлитных клеток EOM могут контролироваться нишей. В поддержку этой гипотезы недавнее исследование показало, что интерстициальные клетки PW1/peg + (PIC), которые присутствуют в большем количестве в EOM по сравнению с TA, обеспечивают промиогенную среду, которая способствует устойчивости как сателлитных клеток EOM и миофибрилл к дистрофическим и возрастным заболеваниям (Formicola et al., 2014). Вместе эти данные поднимают некоторые интригующие вопросы. Какую роль играют внутренние и внешние механизмы в биологии сателлитных клеток EOM? Можно ли манипулировать такими механизмами для улучшения качества жизни людей, страдающих миопатическими заболеваниями? Исследования все еще необходимы для дальнейшего выяснения механизмов, способствующих уникальным фенотипам этих сателлитных клеток.
., 2009). Эти данные свидетельствуют о том, что фенотипы сателлитных клеток EOM могут контролироваться нишей. В поддержку этой гипотезы недавнее исследование показало, что интерстициальные клетки PW1/peg + (PIC), которые присутствуют в большем количестве в EOM по сравнению с TA, обеспечивают промиогенную среду, которая способствует устойчивости как сателлитных клеток EOM и миофибрилл к дистрофическим и возрастным заболеваниям (Formicola et al., 2014). Вместе эти данные поднимают некоторые интригующие вопросы. Какую роль играют внутренние и внешние механизмы в биологии сателлитных клеток EOM? Можно ли манипулировать такими механизмами для улучшения качества жизни людей, страдающих миопатическими заболеваниями? Исследования все еще необходимы для дальнейшего выяснения механизмов, способствующих уникальным фенотипам этих сателлитных клеток.
Сателлитные ячейки Masseter
Взрослые жевательные клетки-сателлиты возникают из первой и второй глоточных дуг параксиальной мезодермы черепа с участием мезодермы внутренних органов и экспрессируют уникальный профиль транскрипции по сравнению с сателлитными клетками задних конечностей (рис. 1; Kelly et al., 2004; Ноден и Фрэнсис-Уэст, 2006 г.; Натан и др., 2008 г.; Самбасиван и др., 2009). Ранние исследования in vivo продемонстрировали нарушение регенеративной способности жевательных мышц по сравнению с мышцами задних конечностей, что было связано с меньшим количеством сателлитных клеток во время регенерации (Pavlath et al., 1998). Сателлитные клетки жевательных мышц подвергаются длительным периодам пролиферации in vitro с одновременной задержкой начала дифференцировки (Ono et al., 2010), что потенциально может способствовать нарушению регенеративного ответа, наблюдаемому при остром повреждении жевательной мышцы.Напротив, жевательные сателлитные клетки увеличиваются в количестве на мышечное волокно с возрастом, в то время как их пролиферативная способность снижается ex vivo (Ono et al., 2010). Какие молекулярные пути связаны с возрастным увеличением числа сателлитных клеток в жевательных мышцах по сравнению с уменьшением числа сателлитных клеток в других скелетных мышцах? Мало что известно о лежащих в основе молекулярных механизмах, управляющих фенотипами жевательных сателлитных клеток.
1; Kelly et al., 2004; Ноден и Фрэнсис-Уэст, 2006 г.; Натан и др., 2008 г.; Самбасиван и др., 2009). Ранние исследования in vivo продемонстрировали нарушение регенеративной способности жевательных мышц по сравнению с мышцами задних конечностей, что было связано с меньшим количеством сателлитных клеток во время регенерации (Pavlath et al., 1998). Сателлитные клетки жевательных мышц подвергаются длительным периодам пролиферации in vitro с одновременной задержкой начала дифференцировки (Ono et al., 2010), что потенциально может способствовать нарушению регенеративного ответа, наблюдаемому при остром повреждении жевательной мышцы.Напротив, жевательные сателлитные клетки увеличиваются в количестве на мышечное волокно с возрастом, в то время как их пролиферативная способность снижается ex vivo (Ono et al., 2010). Какие молекулярные пути связаны с возрастным увеличением числа сателлитных клеток в жевательных мышцах по сравнению с уменьшением числа сателлитных клеток в других скелетных мышцах? Мало что известно о лежащих в основе молекулярных механизмах, управляющих фенотипами жевательных сателлитных клеток.
Фарингеальные сателлитные клетки
Глотание зависит от синхронного сокращения семи основных мышц, выстилающих носовую, ротовую и гортанную части глотки, чтобы обеспечить продвижение пищи и жидкости из ротовой полости в пищевод (Donner et al., 1985; Рубесин и др., 1987; Экберг и др., 2009). Мышцы глотки возникают из краниальной параксиальной мезодермы третьей и четвертой глоточных дуг с участием внутренностной мезодермы (Рисунок 1; Kelly et al., 2004; Noden and Francis-West, 2006; Nathan et al., 2008). Мышцы глотки включают шилоглоточную, небно-глоточную, сальпингофарингеальную, а также верхнюю, среднюю и нижнюю констрикторы глотки (Dutta and Basmajian, 1960; Himmelreich, 1973; Donner et al., 1985; Rubesin et al., 1987; Экберг и др., 2009). Нижний констриктор глотки можно подразделить на перстнеглоточную и щитоглоточную мышцы (Donner et al., 1985; Rubesin et al., 1987; Ekberg et al., 2009). Недавние исследования in vivo и in vitro показали, что сателлитные клетки глотки обладают уникальными биологическими характеристиками по сравнению с сателлитными клетками задних конечностей. Мышцы глотки содержат повышенное количество активированных и пролиферирующих сателлитных клеток и вносят вклад в новые миоядра миофибрилл глотки при отсутствии индуцированного повреждения (Randolph et al., 2015). Кроме того, клональные анализы in vitro предполагают, что высокопролиферативная субпопуляция фарингеальных сателлитных клеток может способствовать пролиферативному фенотипу (Randolph et al., 2015). Исследования абляции in vivo показали, что сателлитные клетки необходимы для поддержания числа миоядер в определенных мышцах глотки в исходных условиях, предполагая, что мышцы глотки подвергаются миоядерному обновлению и нуждаются в миоядерном добавлении в результате продолжающегося миогенеза сателлитных клеток (Randolph et al., 2015). Могут ли патологические мутации усилить миоядерный обмен в мышцах глотки до такой степени, что сателлитные клетки глотки больше не смогут адекватно поставлять достаточное количество миоядер для поддержания гомеостаза? Нарушают ли вызывающие заболевание мутации базальный миогенный фенотип сателлитных клеток глотки, потенциально способствуя патологической чувствительности мышц глотки, наблюдаемой при некоторых MD? Эти и многие другие вопросы еще предстоит решить.
Мышцы глотки содержат повышенное количество активированных и пролиферирующих сателлитных клеток и вносят вклад в новые миоядра миофибрилл глотки при отсутствии индуцированного повреждения (Randolph et al., 2015). Кроме того, клональные анализы in vitro предполагают, что высокопролиферативная субпопуляция фарингеальных сателлитных клеток может способствовать пролиферативному фенотипу (Randolph et al., 2015). Исследования абляции in vivo показали, что сателлитные клетки необходимы для поддержания числа миоядер в определенных мышцах глотки в исходных условиях, предполагая, что мышцы глотки подвергаются миоядерному обновлению и нуждаются в миоядерном добавлении в результате продолжающегося миогенеза сателлитных клеток (Randolph et al., 2015). Могут ли патологические мутации усилить миоядерный обмен в мышцах глотки до такой степени, что сателлитные клетки глотки больше не смогут адекватно поставлять достаточное количество миоядер для поддержания гомеостаза? Нарушают ли вызывающие заболевание мутации базальный миогенный фенотип сателлитных клеток глотки, потенциально способствуя патологической чувствительности мышц глотки, наблюдаемой при некоторых MD? Эти и многие другие вопросы еще предстоит решить. Тем не менее, несколько исследований изучали биологию сателлитных клеток глотки в контексте окулофарингеальной мышечной дистрофии и будут обсуждаться ниже.
Тем не менее, несколько исследований изучали биологию сателлитных клеток глотки в контексте окулофарингеальной мышечной дистрофии и будут обсуждаться ниже.
Может ли биология вариабельной сателлитной клетки способствовать дифференциальной восприимчивости определенных мышц к дистрофическим мутациям?
Мышечные дистрофии представляют собой группу дегенеративных мышечных заболеваний, обусловленных мутациями в белках с различной функциональностью, таких как структура сарколеммы (Hoffman et al., 1987), структура ядерной оболочки (Bione et al., 1994; Bonne et al., 1999; Mittelbronn et al., 2008), посттрансляционное гликозилирование (Bonnemann et al., 2014) и связывание РНК (Brais et al., 1998; Кюн и др., 2009). Многие мутации были охарактеризованы как в специфических для скелетных мышц, так и повсеместно экспрессируемых генах, но обе проявляются в мышечной патологии. Любопытно, что каждая дистрофия поражает определенную группу скелетных мышц человеческого тела (Emery, 2002), что позволяет предположить, что между отдельными мышцами существуют биологические различия, предрасполагающие их к определенным патологическим этиологиям.
Сателлитные клетки вовлечены в патологию некоторых MD и могут способствовать изменчивой мышечной чувствительности, наблюдаемой при некоторых дистрофиях, посредством нескольких механизмов (таблица 1).Например, в ответ на хроническую дегенерацию миофибрилл сателлитные клетки подвергаются множественным циклам регенерации, которые со временем могут «истощать» регенеративные способности популяции сателлитных клеток (Webster and Blau, 1990; Decary et al., 2000; Morgan). и Zammit, 2010; Sacco et al., 2010). Кроме того, повреждение сателлитных клеток может произойти на ранней стадии заболевания, если сателлитные клетки экспрессируют мутантные гены. Ниже мы обсудим функциональные нарушения в сателлитных клетках, наблюдаемые при болезни Дюшенна (Blau et al., 1983; Вебстер и Блау, 1990 г.; Sacco et al., 2010), Emery-Dreifuss MD (Favreau et al., 2004; Frock et al., 2006), facioscapulohumeral MD (Winokur et al., 2003; Barro et al., 2010), миотоническая дистрофия (Furling et al., 2001; Thornell et al. , 2009), окулофарингеальной MD (Périé et al., 2006), а также некоторых врожденных MD (Castets et al., 2011; Urciuolo et al., 2013).
, 2009), окулофарингеальной MD (Périé et al., 2006), а также некоторых врожденных MD (Castets et al., 2011; Urciuolo et al., 2013).
Мышечная дистрофия Дюшенна
Мышечная дистрофия Дюшенна (МДД) — Х-сцепленное заболевание с ранним началом у детей, связанное с отсутствием дистрофина (Hoffman et al., 1987), ассоциированный с сарколеммой цитоплазматический белок, критический для поддержания целостности сарколеммы миофибрилл (Durbeej and Campbell, 2002). Минимальные уровни механического стресса необходимы для нарушения целостности сарколеммы в отсутствие дистрофина, что приводит к повторяющимся раундам повреждения и репарации миофибрилл (Petrof et al., 1993). Пациенты, страдающие МДД, испытывают прогрессирующую потерю мышечной функции, что в конечном итоге приводит к смерти в возрасте до 30 лет. Основные скелетные мышцы, поражаемые при МДД, находятся в плече, верхних конечностях, бедрах, бедрах и икрах (Emery, 2002).Опасные для жизни симптомы у многих пациентов включают сердечную и дыхательную недостаточность из-за поражения мышц сердца и диафрагмы соответственно (Nigro et al. , 1990; Stedman et al., 1991). Следует отметить, что черепно-лицевые мышцы, такие как экстраокулярные и внутренние мышцы гортани, в большинстве случаев не повреждаются при МДД, за исключением мышц глотки на поздних стадиях заболевания (Kaminski et al., 1992; Emery, 2002; Marques et al., 2007). ; Шинонага и др., 2008). Механизм устойчивости ЭОМ к дистрофическим изменениям был недавно рассмотрен в исследованиях облучения с использованием гетерозиготных мышей mdx:utrophin .В ЭСО этих мышей не развивались дистрофические фенотипы даже после гамма-облучения 18 Гр. Эта устойчивость к дистрофическим изменениям была связана с наличием множества популяций миогенных предшественников ЭОМ, которые предотвращали потерю размера миофибрилл, предполагая, что пролиферативные популяции сателлитных клеток ЭОМ играют роль в сохранении мышц ЭОМ при многих дистрофиях (McDonald et al., 2014). Аномалии сателлитных клеток отмечаются в дефицитных по дистрофину мышцах задних конечностей как у мышей, так и у людей на ранних стадиях заболевания.
, 1990; Stedman et al., 1991). Следует отметить, что черепно-лицевые мышцы, такие как экстраокулярные и внутренние мышцы гортани, в большинстве случаев не повреждаются при МДД, за исключением мышц глотки на поздних стадиях заболевания (Kaminski et al., 1992; Emery, 2002; Marques et al., 2007). ; Шинонага и др., 2008). Механизм устойчивости ЭОМ к дистрофическим изменениям был недавно рассмотрен в исследованиях облучения с использованием гетерозиготных мышей mdx:utrophin .В ЭСО этих мышей не развивались дистрофические фенотипы даже после гамма-облучения 18 Гр. Эта устойчивость к дистрофическим изменениям была связана с наличием множества популяций миогенных предшественников ЭОМ, которые предотвращали потерю размера миофибрилл, предполагая, что пролиферативные популяции сателлитных клеток ЭОМ играют роль в сохранении мышц ЭОМ при многих дистрофиях (McDonald et al., 2014). Аномалии сателлитных клеток отмечаются в дефицитных по дистрофину мышцах задних конечностей как у мышей, так и у людей на ранних стадиях заболевания. В мышцах задних конечностей мышей нарушение прикрепления сателлитных клеток к миофибриллам mdx может способствовать гипотрофии миофибрилл до мионекроза, обнаруживаемой при раннем постнатальном заболевании (Duddy et al., 2015). Кроме того, преждевременное старение клеток наблюдается в культивируемых миобластах, выделенных из мышц конечностей пациентов с МДД, уже в возрасте 2 лет, в возрасте типичного клинического начала (Blau et al., 1983; Webster and Blau, 1990), что объясняется дефицитом обоих Передача сигналов Notch и укорочение теломер после повторяющихся регенеративных циклов (Mouly et al., 2005; Сакко и др., 2010 г.; Цзян и др., 2014).
В мышцах задних конечностей мышей нарушение прикрепления сателлитных клеток к миофибриллам mdx может способствовать гипотрофии миофибрилл до мионекроза, обнаруживаемой при раннем постнатальном заболевании (Duddy et al., 2015). Кроме того, преждевременное старение клеток наблюдается в культивируемых миобластах, выделенных из мышц конечностей пациентов с МДД, уже в возрасте 2 лет, в возрасте типичного клинического начала (Blau et al., 1983; Webster and Blau, 1990), что объясняется дефицитом обоих Передача сигналов Notch и укорочение теломер после повторяющихся регенеративных циклов (Mouly et al., 2005; Сакко и др., 2010 г.; Цзян и др., 2014).
Поясно-конечностная мышечная дистрофия
Поясно-конечностные мышечные дистрофии (LGMD) связаны с мутациями более 20 различных генов как в мышечно-специфических, так и повсеместно экспрессируемых генах с рядом молекулярных функций (Vieira et al., 2014). К ним относятся белки саркомера (титин), белки сарколеммы (саркогликан), гликозилтрансферазы (фукутин), белки ядерной оболочки (ламин A/C) и белки процессинга РНК (HNRPDL). Интересно, что, несмотря на огромное этиологическое разнообразие, все мутации вызывают дистрофические изменения в мышцах верхней конечности, плеча, груди, бедра и бедра (Broglio et al., 2010; Mitsuhashi and Kang, 2012). Вовлечение сателлитных клеток было связано с некоторыми LGMD. Биопсии пациентов с LGMD2A продемонстрировали снижение miR-1 и miR206, микроРНК, участвующих в облегчении дифференцировки сателлитных клеток, что коррелирует с увеличением популяции Pax7+. Несмотря на увеличение популяции Pax7+, регенерация была нарушена, а фиброз повышен, что позволяет предположить, что нарушение перехода сателлитных клеток от пролиферации к дифференцировке может способствовать патологии LGMD2A (Rosales et al., 2013). Напротив, у пациентов с мутациями α-, β- или γ-саркогликанов (LGMD2D, 2E и 2C соответственно) сообщалось о снижении числа сателлитных клеток по сравнению с образцами пациентов с мышечной дистрофией Беккера (Higuchi et al., 1999). Кроме того, мышиные POMGnT1-нулевые миобласты (представители LGMD2O) продемонстрировали нарушение пролиферации in vitro (Yoshida et al.
Интересно, что, несмотря на огромное этиологическое разнообразие, все мутации вызывают дистрофические изменения в мышцах верхней конечности, плеча, груди, бедра и бедра (Broglio et al., 2010; Mitsuhashi and Kang, 2012). Вовлечение сателлитных клеток было связано с некоторыми LGMD. Биопсии пациентов с LGMD2A продемонстрировали снижение miR-1 и miR206, микроРНК, участвующих в облегчении дифференцировки сателлитных клеток, что коррелирует с увеличением популяции Pax7+. Несмотря на увеличение популяции Pax7+, регенерация была нарушена, а фиброз повышен, что позволяет предположить, что нарушение перехода сателлитных клеток от пролиферации к дифференцировке может способствовать патологии LGMD2A (Rosales et al., 2013). Напротив, у пациентов с мутациями α-, β- или γ-саркогликанов (LGMD2D, 2E и 2C соответственно) сообщалось о снижении числа сателлитных клеток по сравнению с образцами пациентов с мышечной дистрофией Беккера (Higuchi et al., 1999). Кроме того, мышиные POMGnT1-нулевые миобласты (представители LGMD2O) продемонстрировали нарушение пролиферации in vitro (Yoshida et al. , 2001; Miyagoe-Suzuki et al., 2009). В мышиной модели LGMD2H нокаут E3-убиквитинлигазы, содержащей трехкомпонентный мотив 32 (TRIM32), приводил к старению сателлитных клеток как in vitro , так и in vivo (Kudryashova et al., 2012; Мохонова и др., 2015). Интересно, что фармакологическая индукция экспрессии фоллистатина в сателлитных клетках с использованием ингибитора деацетилазы, трихостатина А, оказалась благоприятной для восстановления размера миофибрилл в α-sarcoglycan-дефицитных LGMD in vivo (Minetti et al., 2006). Следует отметить, что обработка in vitro сателлитных клеток мышей с дефицитом α-саркогликанов трихостатином А приводила к гипернуклеации мышечных трубок, что свидетельствует о фармакологическом усилении дифференцировки/слияния миобластов (Minetti et al., 2006). Еще неизвестно, могут ли другие фармакологические подходы, которые изменяют функцию сателлитных клеток, также принести пользу пациентам с LGMD с другими мутациями.
, 2001; Miyagoe-Suzuki et al., 2009). В мышиной модели LGMD2H нокаут E3-убиквитинлигазы, содержащей трехкомпонентный мотив 32 (TRIM32), приводил к старению сателлитных клеток как in vitro , так и in vivo (Kudryashova et al., 2012; Мохонова и др., 2015). Интересно, что фармакологическая индукция экспрессии фоллистатина в сателлитных клетках с использованием ингибитора деацетилазы, трихостатина А, оказалась благоприятной для восстановления размера миофибрилл в α-sarcoglycan-дефицитных LGMD in vivo (Minetti et al., 2006). Следует отметить, что обработка in vitro сателлитных клеток мышей с дефицитом α-саркогликанов трихостатином А приводила к гипернуклеации мышечных трубок, что свидетельствует о фармакологическом усилении дифференцировки/слияния миобластов (Minetti et al., 2006). Еще неизвестно, могут ли другие фармакологические подходы, которые изменяют функцию сателлитных клеток, также принести пользу пациентам с LGMD с другими мутациями.
Мышечная дистрофия Эмери-Дрейфуса
Мышечная дистрофия Эмери-Дрейфуса приводит к прогрессирующей слабости плечевых, верхних конечностей и икроножных мышц пациентов. Наиболее распространенные формы этой дистрофии вызываются мутациями повсеместно экспрессируемых белков ядерной оболочки эмерина, ламина А или ламина С (Helbling-Leclerc et al., 2002). Почему при этом заболевании преимущественно поражаются скелетные и сердечные мышцы, до сих пор неясно. Пациенты с МД Эмери-Дрейфуса могут иметь тяжелую сердечную патологию уже в возрасте 30 лет (Vohanka et al., 2001; Emery, 2002; Broglio et al., 2010). В исследованиях in vitro с использованием первичных мышечных культур нокаутных мышей Lmna -/- , сверхэкспрессии мутантного ламина A R453W или нокдауна РНК-интерференции emerin были обнаружены дефекты дифференцировки миобластов (Favreau et al., 2004; Фрок и др., 2006). В недавнем исследовании in vitro культура миобластов, полученных от пациентов, лишенных эмерина, продемонстрировала усиленную пролиферацию со спонтанной дифференцировкой по сравнению с контрольными миобластами, что позволяет предположить, что нарушение сателлитных клеток может играть роль в болезни Эмери-Дрейфуса (Meinke et al.
Наиболее распространенные формы этой дистрофии вызываются мутациями повсеместно экспрессируемых белков ядерной оболочки эмерина, ламина А или ламина С (Helbling-Leclerc et al., 2002). Почему при этом заболевании преимущественно поражаются скелетные и сердечные мышцы, до сих пор неясно. Пациенты с МД Эмери-Дрейфуса могут иметь тяжелую сердечную патологию уже в возрасте 30 лет (Vohanka et al., 2001; Emery, 2002; Broglio et al., 2010). В исследованиях in vitro с использованием первичных мышечных культур нокаутных мышей Lmna -/- , сверхэкспрессии мутантного ламина A R453W или нокдауна РНК-интерференции emerin были обнаружены дефекты дифференцировки миобластов (Favreau et al., 2004; Фрок и др., 2006). В недавнем исследовании in vitro культура миобластов, полученных от пациентов, лишенных эмерина, продемонстрировала усиленную пролиферацию со спонтанной дифференцировкой по сравнению с контрольными миобластами, что позволяет предположить, что нарушение сателлитных клеток может играть роль в болезни Эмери-Дрейфуса (Meinke et al. , 2015).
, 2015).
Лицево-лопаточно-плечевая мышечная дистрофия
Лицево-лопаточно-плечевая мышечная дистрофия (FSHD) названа в честь мышц, в основном поражаемых при заболевании, лицевых, плечевых мышц и мышц плеча, но могут также поражаться мышцы стопы и тазобедренного пояса (Tawil and Van Der Maarel, 2006).Из дистрофий, поражающих черепно-лицевые мышцы, ЛЛПД дает наилучший прогноз для долгосрочного выживания, так как это медленно прогрессирующее заболевание, редко поражающее сердце или способность дышать (Tawil and Van Der Maarel, 2006). Причинная делеция для FSHD типа 1 (FSHD1) происходит в субтеломерной области хромосомы 4, что может индуцировать экспрессию генов, таких как ген 1 области FSHD ( FGR1 ), FGR2 , ANT1 , DUX4. и DUX4c (Gabellini et al., 2002; Диксит и др., 2007 г.; Анссо и др., 2009 г.; Бодега и др., 2009 г.; Снайдер и др., 2010). Патогенный вклад этих генов в ЛЛПД все еще изучается. Однако данных об участии сателлитных клеток в FSHD1 становится все больше. Экспрессия DUX4 в культивируемых миобластах ингибировала миогенную дифференцировку за счет репрессии Myf5 и MyoD (Bosnakovski et al., 2008a,b, 2009), в то время как сверхэкспрессия DUX4 была токсична для миобластов in vitro 9021 (Kowal), 2007). Напротив, сверхэкспрессия DUX4c стимулирует пролиферацию миобластов, но ингибирует дифференцировку in vitro (Bosnakovski et al., 2008a; Ansseau et al., 2009). Кроме того, сверхэкспрессия FGR1 нарушает пролиферацию миобластов, а также слияние миобластов (Chen et al., 2011; Feeney et al., 2015). Первичные миобласты, собранные из пораженных мышц бедра трансгенных мышей со сверхэкспрессией FGR1, продуцировали клональные колонии меньшего размера, чем миобласты, полученные из непораженной мышцы диафрагмы (Chen et al., 2011). В совокупности эти результаты показывают, что сателлитные клетки могут играть непосредственную роль в патологии FSHD1. Механизм(ы), лежащий в основе мышечной специфичности этих измененных миогенных фенотипов, еще предстоит определить.
Экспрессия DUX4 в культивируемых миобластах ингибировала миогенную дифференцировку за счет репрессии Myf5 и MyoD (Bosnakovski et al., 2008a,b, 2009), в то время как сверхэкспрессия DUX4 была токсична для миобластов in vitro 9021 (Kowal), 2007). Напротив, сверхэкспрессия DUX4c стимулирует пролиферацию миобластов, но ингибирует дифференцировку in vitro (Bosnakovski et al., 2008a; Ansseau et al., 2009). Кроме того, сверхэкспрессия FGR1 нарушает пролиферацию миобластов, а также слияние миобластов (Chen et al., 2011; Feeney et al., 2015). Первичные миобласты, собранные из пораженных мышц бедра трансгенных мышей со сверхэкспрессией FGR1, продуцировали клональные колонии меньшего размера, чем миобласты, полученные из непораженной мышцы диафрагмы (Chen et al., 2011). В совокупности эти результаты показывают, что сателлитные клетки могут играть непосредственную роль в патологии FSHD1. Механизм(ы), лежащий в основе мышечной специфичности этих измененных миогенных фенотипов, еще предстоит определить.
Миотоническая дистрофия
Миотоническая дистрофия (DM) представляет собой сложную мультисистемную группу дистрофий, которые генетически возникают из-за экспансии нетранслируемых повторяющихся нуклеотидов двух отдельных генов, миотонической протеинкиназы дистофии ( DMPK ) и белка цинковых пальцев 9 ( ZNF9 ) (Дэй и Ранум, 2005).Повтор (CTG) 80-4000 в 3′-нетранслируемой области DMPK присутствует у пациентов с миотонической дистрофией 1 типа (СД1). Расширенные области транскриптов DMPK приводят к изменению биогенеза РНК и процессинга множественных транскриптов, частично за счет секвестрации фактора сплайсинга мышечной слепоты (MBNL1) и стабилизации CUG-связывающего белка 1 (CUGBP1; Mastroyiannopoulos et al., 2010). При миотонической дистрофии 2 типа (СД2) до 75–11 000 повторных экспансий (TG) n (TCTG) n (CCTG) n находятся в интроне 1 ZNF9 (Day and Ranum, 2005). , нарушение регуляции альтернативного слайсинга, а также продукции белка за счет секвестрации протеасомы 20S (Salisbury et al., 2009). В то время как СД 1 и СД 2 возникают в результате различных генетических мутаций, биологические последствия аналогичны: миотония, мышечная дистрофия, мышечные боли, катаракта, сердечные аритмии, нечувствительность к инсулину, диабет, гипогаммаглобулинемия и тестикулярная недостаточность возникают в обоих случаях (Schoser and Timchenko, 2010). . СД поражает мышцы век, лица, шеи, предплечий и ног, диафрагму и межреберные мышцы (Batten, Gibb, 1909; Zifko et al., 1996). Тем не менее, СД1 связан с мышечной слабостью и атрофией мышц нижних конечностей, в то время как при СД2 заболевание чаще поражает верхние конечности (Tieleman et al., 2012). Могут возникнуть опасные для жизни состояния, включающие сердечные заболевания, дыхательную недостаточность и трудности при глотании (Zifko et al., 1996; Tieleman et al., 2009, 2012). Количество сателлитных клеток, их пролиферация и дифференцировка дифференцированно изменены у больных СД1.
, нарушение регуляции альтернативного слайсинга, а также продукции белка за счет секвестрации протеасомы 20S (Salisbury et al., 2009). В то время как СД 1 и СД 2 возникают в результате различных генетических мутаций, биологические последствия аналогичны: миотония, мышечная дистрофия, мышечные боли, катаракта, сердечные аритмии, нечувствительность к инсулину, диабет, гипогаммаглобулинемия и тестикулярная недостаточность возникают в обоих случаях (Schoser and Timchenko, 2010). . СД поражает мышцы век, лица, шеи, предплечий и ног, диафрагму и межреберные мышцы (Batten, Gibb, 1909; Zifko et al., 1996). Тем не менее, СД1 связан с мышечной слабостью и атрофией мышц нижних конечностей, в то время как при СД2 заболевание чаще поражает верхние конечности (Tieleman et al., 2012). Могут возникнуть опасные для жизни состояния, включающие сердечные заболевания, дыхательную недостаточность и трудности при глотании (Zifko et al., 1996; Tieleman et al., 2009, 2012). Количество сателлитных клеток, их пролиферация и дифференцировка дифференцированно изменены у больных СД1. Снижение числа сателлитных клеток может быть результатом индуцированных аутофагических процессов в миобластах DM1 (Beffy et al., 2010). Культивируемые миобласты, полученные из пораженных мышц нижних конечностей, пролиферировали меньше по сравнению с культурами непораженных мышц верхних конечностей, полученных от тех же пациентов (Thornell et al., 2009). Повышенная экспрессия простагландина E2 миобластами DM1 ингибирует дифференцировку и слияние аутокринным образом (Beaulieu et al., 2012). Кроме того, p16-путь индуцирует преждевременное старение в миобластах DM1 (Bigot et al., 2009). Сателлитные клетки также вовлечены в патологию DM2. Например, сателлитные клетки претерпевают преждевременное старение у пациентов с СД2, но не зависимым от p16 образом посредством теломер-управляемого пути (Malatesta et al., 2011; Rena et al., 2014). Нарушение сателлитных клеток может быть основной патологической детерминантой миотонических дистрофий.
Снижение числа сателлитных клеток может быть результатом индуцированных аутофагических процессов в миобластах DM1 (Beffy et al., 2010). Культивируемые миобласты, полученные из пораженных мышц нижних конечностей, пролиферировали меньше по сравнению с культурами непораженных мышц верхних конечностей, полученных от тех же пациентов (Thornell et al., 2009). Повышенная экспрессия простагландина E2 миобластами DM1 ингибирует дифференцировку и слияние аутокринным образом (Beaulieu et al., 2012). Кроме того, p16-путь индуцирует преждевременное старение в миобластах DM1 (Bigot et al., 2009). Сателлитные клетки также вовлечены в патологию DM2. Например, сателлитные клетки претерпевают преждевременное старение у пациентов с СД2, но не зависимым от p16 образом посредством теломер-управляемого пути (Malatesta et al., 2011; Rena et al., 2014). Нарушение сателлитных клеток может быть основной патологической детерминантой миотонических дистрофий.
Окулофарингеальная мышечная дистрофия
Окулофарингеальная мышечная дистрофия (OPMD) является аутосомно-доминантным заболеванием, которое обычно поражает людей старше 50 лет (Abu-Baker and Rouleau, 2007; Messaed and Rouleau, 2009). Аберрантная экспансия аланинов на N-конце полиаденозин-связывающего ядерного белка (PABPN1) является основной причиной этого в настоящее время неизлечимого заболевания (Brais et al., 1998; Abu-Baker and Rouleau, 2007; Messaed and Rouleau, 2009). ). PABPN1 — повсеместно экспрессируемый белок, играющий ключевую роль в биогенезе РНК (Banerjee et al., 2013). Эндогенный белок содержит 10-аланиновый повтор на своем N-конце, но у пациентов с OPMD сообщается об экспансии, приводящей к 12–18 аланинам (Brais et al., 1998; Abu-Baker and Rouleau, 2007; Messaed and Rouleau, 2009; Джоуан и др., 2014). Мышечная слабость и дистрофия возникают преимущественно в черепно-лицевых скелетных мышцах, включая верхнее веко, глотку, ЭСО и язык со слабостью в мышцах верхних конечностей, развивающейся позже при заболевании (Emery, 2002; Abu-Baker and Rouleau, 2007; Messaed and Rouleau). , 2009).
Аберрантная экспансия аланинов на N-конце полиаденозин-связывающего ядерного белка (PABPN1) является основной причиной этого в настоящее время неизлечимого заболевания (Brais et al., 1998; Abu-Baker and Rouleau, 2007; Messaed and Rouleau, 2009). ). PABPN1 — повсеместно экспрессируемый белок, играющий ключевую роль в биогенезе РНК (Banerjee et al., 2013). Эндогенный белок содержит 10-аланиновый повтор на своем N-конце, но у пациентов с OPMD сообщается об экспансии, приводящей к 12–18 аланинам (Brais et al., 1998; Abu-Baker and Rouleau, 2007; Messaed and Rouleau, 2009; Джоуан и др., 2014). Мышечная слабость и дистрофия возникают преимущественно в черепно-лицевых скелетных мышцах, включая верхнее веко, глотку, ЭСО и язык со слабостью в мышцах верхних конечностей, развивающейся позже при заболевании (Emery, 2002; Abu-Baker and Rouleau, 2007; Messaed and Rouleau). , 2009).
Главной опасной для жизни проблемой для пациентов с OPMD является возникающая в результате дисфагия или нарушение глотания (Périé et al. , 2006). Глоточные мышцы носовой, ротовой и гортанной глотки являются важнейшими компонентами глотательного рефлекса (Миллер, 2002; Эртекин, Айдогду, 2003; Миллер, 2008), который предотвращает аспирацию пищи и воды в трахею и легкие и образование опасная для жизни пневмония (Martin et al., 1994; Prasse and Kikano, 2009). Следует отметить, что снижение пролиферации сателлитных клеток глотки, выделенных у пациентов с OPMD, наблюдалось in vitro (Périé et al., 2006). У трансгенных мышей, которые сверхэкспрессируют PABPN1 дикого типа , в частности, в скелетных мышцах, увеличенное количество миофибрилл с центральными ядрами указывает на положительный эффект PABPN1 дикого типа на слияние сателлитных клеток in vivo (Randolph et al., 2014). В совокупности вышеупомянутые исследования показывают, что PABPN1 играет критическую роль в миогенезе сателлитных клеток глотки, и мутации в этом белке могут способствовать нарушению сателлитных клеток у пациентов с OPMD.
, 2006). Глоточные мышцы носовой, ротовой и гортанной глотки являются важнейшими компонентами глотательного рефлекса (Миллер, 2002; Эртекин, Айдогду, 2003; Миллер, 2008), который предотвращает аспирацию пищи и воды в трахею и легкие и образование опасная для жизни пневмония (Martin et al., 1994; Prasse and Kikano, 2009). Следует отметить, что снижение пролиферации сателлитных клеток глотки, выделенных у пациентов с OPMD, наблюдалось in vitro (Périé et al., 2006). У трансгенных мышей, которые сверхэкспрессируют PABPN1 дикого типа , в частности, в скелетных мышцах, увеличенное количество миофибрилл с центральными ядрами указывает на положительный эффект PABPN1 дикого типа на слияние сателлитных клеток in vivo (Randolph et al., 2014). В совокупности вышеупомянутые исследования показывают, что PABPN1 играет критическую роль в миогенезе сателлитных клеток глотки, и мутации в этом белке могут способствовать нарушению сателлитных клеток у пациентов с OPMD.
Результаты недавних клинических испытаний предоставляют предварительные доказательства использования трансплантации сателлитных клеток в качестве терапевтического лечения пациентов с дисфагией OPMD. Клинические испытания фазы I/IIa проводились с участием пациентов с дисфагией OPMD, у которых миобласты, полученные из непораженных скелетных мышц, амплифицировали в культуре и трансплантировали в перстнеглоточные мышцы после хирургического лечения перстнеглоточной мышцы. Пациенты, получавшие инъекции большего количества миобластов в крикофарингеальную область, продемонстрировали значительное улучшение глотания в течение 2-летнего периода (Perie et al., 2014), что обеспечило экспериментальную поддержку использования терапии на основе сателлитных клеток для пациентов с OPMD.
Клинические испытания фазы I/IIa проводились с участием пациентов с дисфагией OPMD, у которых миобласты, полученные из непораженных скелетных мышц, амплифицировали в культуре и трансплантировали в перстнеглоточные мышцы после хирургического лечения перстнеглоточной мышцы. Пациенты, получавшие инъекции большего количества миобластов в крикофарингеальную область, продемонстрировали значительное улучшение глотания в течение 2-летнего периода (Perie et al., 2014), что обеспечило экспериментальную поддержку использования терапии на основе сателлитных клеток для пациентов с OPMD.
Врожденные мышечные дистрофии
Врожденные мышечные дистрофии (ВМД) представляют собой большую группу врожденных мышечных заболеваний. Хотя эта группа мышечных заболеваний широко изучена, на сегодняшний день участие сателлитных клеток вовлечено в патологию только двух форм: миопатий, связанных с коллагеном VI (COL6-RD), и миопатий, связанных с селенопротеином N (SEPN1-RM). COL6-RD включает болезнь Ульриха и миопатию Бетлема и возникает в результате мутаций в COL6A1, COL6A2 или COL6A3 (Bonnemann et al. , 2014). Мышечная слабость возникает в дистальных отделах конечностей, мышцах-сгибателях шеи, поясничных мышцах, межреберных мышцах и диафрагме, а также в мышцах проксимальных отделов конечностей, таких как четырехглавая, бицепс и трицепс (Haq et al., 1999; Camacho Vanegas et al., 2001; Quijano- Рой и др., 2012). Когда образцы биопсии человека исследовали на предмет изменения количества сателлитных клеток Pax7 + в норме по сравнению с Ullrich MD, никаких изменений не наблюдалось (Paco et al., 2012). Однако у мышей с нокаутом Col6a1 самообновление сателлитных клеток было нарушено после нескольких эпизодов индуцированного повреждения, в то время как количество сателлитных клеток сохранялось у мышей Col6a1 -/- при лечении циклоспорином А (Urciuolo et al., 2013; Гаттаццо и др., 2014). Еще предстоит определить, повторяются ли эти результаты в сателлитных клетках пациентов с МД Ульриха. Синдром ригидного позвоночника, связанный с SEPN1 , является CMD в рамках миопатий, связанных с SEPN1.
, 2014). Мышечная слабость возникает в дистальных отделах конечностей, мышцах-сгибателях шеи, поясничных мышцах, межреберных мышцах и диафрагме, а также в мышцах проксимальных отделов конечностей, таких как четырехглавая, бицепс и трицепс (Haq et al., 1999; Camacho Vanegas et al., 2001; Quijano- Рой и др., 2012). Когда образцы биопсии человека исследовали на предмет изменения количества сателлитных клеток Pax7 + в норме по сравнению с Ullrich MD, никаких изменений не наблюдалось (Paco et al., 2012). Однако у мышей с нокаутом Col6a1 самообновление сателлитных клеток было нарушено после нескольких эпизодов индуцированного повреждения, в то время как количество сателлитных клеток сохранялось у мышей Col6a1 -/- при лечении циклоспорином А (Urciuolo et al., 2013; Гаттаццо и др., 2014). Еще предстоит определить, повторяются ли эти результаты в сателлитных клетках пациентов с МД Ульриха. Синдром ригидного позвоночника, связанный с SEPN1 , является CMD в рамках миопатий, связанных с SEPN1. Пораженные мышцы включают бедро, большую ягодичную мышцу, паравертебральные, межреберные и грудино-ключично-сосцевидные мышцы (Quijano-Roy et al., 2012). Исследования, изучающие сателлитные клетки у мышей Sepn1-/-, выявили снижение количества сателлитных клеток, нарушение самообновления, усиленную пролиферацию сателлитных клеток и истощение пула сателлитных клеток после одного цикла регенерации (Castets et al., 2011). Эти исследования предполагают потенциальную роль сателлитных клеток в патологии некоторых ВМД. Необходимы дальнейшие исследования, чтобы определить, будут ли сателлитные клетки полезными терапевтическими мишенями для пациентов с CMD.
Пораженные мышцы включают бедро, большую ягодичную мышцу, паравертебральные, межреберные и грудино-ключично-сосцевидные мышцы (Quijano-Roy et al., 2012). Исследования, изучающие сателлитные клетки у мышей Sepn1-/-, выявили снижение количества сателлитных клеток, нарушение самообновления, усиленную пролиферацию сателлитных клеток и истощение пула сателлитных клеток после одного цикла регенерации (Castets et al., 2011). Эти исследования предполагают потенциальную роль сателлитных клеток в патологии некоторых ВМД. Необходимы дальнейшие исследования, чтобы определить, будут ли сателлитные клетки полезными терапевтическими мишенями для пациентов с CMD.
Резюме
Скелетные мышцы представляют собой очень разнообразную и динамичную группу тканей. Как обсуждалось, многие факторы способствуют разнообразию скелетных мышц, включая эмбриологическое происхождение, экспрессию генов и функциональные/метаболические потребности. Такое разнообразие, вероятно, способствует патологической чувствительности различных скелетных мышц к старению и болезням. К сожалению, мало что известно о влиянии возраста или заболевания на мышцы, не являющиеся конечностями, в целом или о том, какие факторы предрасполагают их к влиянию патологических состояний. Кроме того, сателлитные клетки могут служить патологическими детерминантами при некоторых дистрофиях; однако наши знания о сателлитных клетках, не относящихся к конечностям, и их роли в биологии мышц крайне недостаточны. Распознавание и выяснение явных различий в биологии сателлитных клеток между разными скелетными мышцами может быть ключом к разгадке загадки специфичности мышц между различными MD.В этом обзоре подчеркивается потенциальная польза изучения биологии сателлитных клеток скелетных мышц, отличных от конечностей, для разработки новых терапевтических подходов к пациентам, страдающим МД.
К сожалению, мало что известно о влиянии возраста или заболевания на мышцы, не являющиеся конечностями, в целом или о том, какие факторы предрасполагают их к влиянию патологических состояний. Кроме того, сателлитные клетки могут служить патологическими детерминантами при некоторых дистрофиях; однако наши знания о сателлитных клетках, не относящихся к конечностям, и их роли в биологии мышц крайне недостаточны. Распознавание и выяснение явных различий в биологии сателлитных клеток между разными скелетными мышцами может быть ключом к разгадке загадки специфичности мышц между различными MD.В этом обзоре подчеркивается потенциальная польза изучения биологии сателлитных клеток скелетных мышц, отличных от конечностей, для разработки новых терапевтических подходов к пациентам, страдающим МД.
Вклад авторов
MR: Концепция и дизайн, финансовая поддержка, сбор и сбор данных, написание рукописи, окончательное утверждение рукописи. GP: Концепция и дизайн, финансовая поддержка, административная поддержка, сбор и сборка данных, написание рукописи, окончательное утверждение рукописи.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Благодарности
Эта работа была поддержана грантами Национального института здравоохранения AR061987 (GP) и DC012225 (MR).
Каталожные номера
Абу-Бакер, А., и Руло, Г.А. (2007). Окулофарингеальная мышечная дистрофия: последние достижения в понимании молекулярных патогенных механизмов и стратегий лечения. Биохим. Биофиз. Acta 1772, 173–185. doi: 10.1016/j.bbadis.2006.10.003
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Анссо, Э., Лаудж-Шенивесс, Д., Марковиц, А., Тассин, А., Вандерпланк, К., Соваж, С., и соавт. (2009). DUX4c активируется при FSHD. Он индуцирует белок MYF5 и пролиферацию миобластов человека. PLoS ONE 4:e7482. doi: 10.1371/journal.pone.0007482
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Аппони, Л. Х., Леунг, С.В., Уильямс, К.Р., Валентини, С.Р., Корбетт, А.Х., и Павлат, Г.К. (2010). Потеря ядерного поли(А)-связывающего белка 1 вызывает дефекты миогенеза и биогенеза мРНК. Гул. Мол. Жене. 19, 1058–1065. doi: 10.1093/hmg/ddp569
Х., Леунг, С.В., Уильямс, К.Р., Валентини, С.Р., Корбетт, А.Х., и Павлат, Г.К. (2010). Потеря ядерного поли(А)-связывающего белка 1 вызывает дефекты миогенеза и биогенеза мРНК. Гул. Мол. Жене. 19, 1058–1065. doi: 10.1093/hmg/ddp569
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Бабюк Р.П., Чжан В., Клагстон Р., Аллан Д.В. и Грир Дж.Дж. (2003). Эмбриологическое происхождение и развитие диафрагмы крыс. Дж. Комп. Нейрол. 455, 477–487. doi: 10.1002/cne.10503
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Барро М., Карнак Г., Флавье С., Мерсье Дж., Васецки Ю. и Лаудж-Шенивес Д. (2010). Миобласты из пораженных и непораженных мышц FSHD демонстрируют дефекты морфологической дифференцировки. Дж. Сотовый. Мол. Мед. 14, 275–289. doi: 10.1111/j.1582-4934.2008.00368.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Бошан, Дж.Р., Хеслоп Л., Ю Д. С. , Тайбахш С., Келли Р. Г., Верниг А. и соавт. (2000). Экспрессия CD34 и Myf5 определяет большинство покоящихся сателлитных клеток взрослых скелетных мышц. J. Cell Biol. 151, 1221–1234. doi: 10.1083/jcb.151.6.1221
, Тайбахш С., Келли Р. Г., Верниг А. и соавт. (2000). Экспрессия CD34 и Myf5 определяет большинство покоящихся сателлитных клеток взрослых скелетных мышц. J. Cell Biol. 151, 1221–1234. doi: 10.1083/jcb.151.6.1221
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Больё, Д., Тебо, П., Пеллетье, Р., Шапделейн, П., Тарнопольски, М., Ферлинг, Д., и другие. (2012). Аномальная продукция простагландина Е2 блокирует миогенную дифференцировку при миотонической дистрофии. Нейробиол. Дис. 45, 122–129. doi: 10.1016/j.nbd.2011.06.014
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Беффи П., Дель Карраторе Р., Масини М., Ферлинг Д., Пуймират Дж., Масиелло П. и др. (2010). Измененные пути передачи сигнала и индукция аутофагии в миобластах человека с миотонической дистрофией 1 типа. Междунар. Дж. Биохим. Клеточная биол. 42, 1973–1983 гг. doi: 10.1016/j.biocel.2010.08.010
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Биго, А. , Кляйн А.Ф., Гаснье Э., Жакмен В., Равассар П., Батлер-Браун Г. и соавт. (2009). Большие повторы CTG запускают p16-зависимое преждевременное старение в клетках-предшественниках миотонической дистрофии 1 типа. утра. Дж. Патол. 174, 1435–1442. doi: 10.2353/ajpath.2009.080560
, Кляйн А.Ф., Гаснье Э., Жакмен В., Равассар П., Батлер-Браун Г. и соавт. (2009). Большие повторы CTG запускают p16-зависимое преждевременное старение в клетках-предшественниках миотонической дистрофии 1 типа. утра. Дж. Патол. 174, 1435–1442. doi: 10.2353/ajpath.2009.080560
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Бионе С., Маэстрини Э., Ривелла С., Манчини М., Реджис С., Ромео Г. и др. (1994). Идентификация нового сцепленного с Х-хромосомой гена, ответственного за мышечную дистрофию Эмери-Дрейфуса. Нац. Жене. 8, 323–327. doi: 10.1038/ng1294-323
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Blanco-Bose, W.E., Yao, C.C., Kramer, R.H., and Blau, H.M. (2001). Очистка первичных миобластов мыши на основе экспрессии альфа-7 интегрина. Экспл. Сотовый рез. 265, 212–220. doi: 10.1006/excr.2001.5191
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Блау, Х. М., Вебстер, К., и Павлат, Г.К. (1983).Дефектные миобласты выявлены при мышечной дистрофии Дюшенна. Проц. Натл. акад. науч. США 80, 4856–4860. doi: 10.1073/pnas.80.15.4856
М., Вебстер, К., и Павлат, Г.К. (1983).Дефектные миобласты выявлены при мышечной дистрофии Дюшенна. Проц. Натл. акад. науч. США 80, 4856–4860. doi: 10.1073/pnas.80.15.4856
Полнотекстовая перекрестная ссылка | Академия Google
Bodega, B., Ramirez, G.D., Grasser, F., Cheli, S., Brunelli, S., Mora, M., et al. (2009). Ремоделирование структуры хроматина локуса лице-лопаточно-плечевой мышечной дистрофии (FSHD) и повышение экспрессии связанного с FSHD гена 1 (FRG1) во время миогенной дифференцировки человека. БМС Биол. 7:41. дои: 10.1186/1741-7007-7-41
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Bonne, G., Di Barletta, M.R., Varnous, S., Becane, H.M., Hammouda, E.H., Merlini, L., et al. (1999). Мутации в гене, кодирующем ламин А/С, вызывают аутосомно-доминантную мышечную дистрофию Эмери-Дрейфуса. Нац. Жене. 21, 285–288. дои: 10.1038/6799
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Боннеманн, К. Г., Ван С. Х., Кихано-Рой С., Деконинк Н., Бертини Э., Феррейро А. и соавт. (2014). Диагностический подход к врожденным мышечным дистрофиям. Нервно-мышечная. Беспорядок. 24, 289–311. doi: 10.1016/j.nmd.2013.12.011
Г., Ван С. Х., Кихано-Рой С., Деконинк Н., Бертини Э., Феррейро А. и соавт. (2014). Диагностический подход к врожденным мышечным дистрофиям. Нервно-мышечная. Беспорядок. 24, 289–311. doi: 10.1016/j.nmd.2013.12.011
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Borello, U., Berarducci, B., Murphy, P., Bajard, L., Buffa, V., Piccolo, S., et al. (2006). Путь Wnt/beta-catenin регулирует Gli-опосредованную экспрессию Myf5 во время сомитогенеза. Разработка 133, 3723–3732.doi: 10.1242/dev.02517
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Борицкий А.Г., Бранк Б., Тайбахш С., Букингем М., Чанг С. и Эмерсон С.П. мл. (1999). Sonic hedgehog контролирует определение эпаксиальных мышц посредством активации Myf5. Разработка 126, 4053–4063.
Академия Google
Боснаковски, Д., Дочери, Р. С., Сюй, З., Слэк, Дж. М., и Киба, М. (2009). Двухфазный миопатический фенотип мышиного DUX, ORF в консервативных повторах, связанных с FSHD. PLoS ONE 4:e7003. doi: 10.1371/journal.pone.0007003
PLoS ONE 4:e7003. doi: 10.1371/journal.pone.0007003
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Боснаковски Д., Лэмб С., Симсек Т., Сюй З., Беляев А., Перлингейро Р. и соавт. (2008а). DUX4c, ген-кандидат ЛЛПД, взаимодействует с регуляторами миогенеза и отменяет дифференцировку миобластов. Экспл. Нейрол. 214, 87–96. doi: 10.1016/j.expneurol.2008.07.022
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Боснаковски, Д., Xu, Z., Gang, E.J., Galindo, C.L., Liu, M., Simsek, T., et al. (2008б). Изогенетический скрининг экспрессии миобластов выявляет опосредованные DUX4 молекулярные патологии, связанные с FSHD. EMBO J. 27, 2766–2779. doi: 10.1038/emboj.2008.201
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Brais, B., Bouchard, J.P., Xie, Y.G., Rochefort, D.L., Chrétien, N., Tomé, F.M., et al. (1998). Короткие экспансии GCG в гене PABP2 вызывают окулофарингеальную мышечную дистрофию. Нац. Жене. 18, 164–167. doi: 10.1038/ng0298-164
Нац. Жене. 18, 164–167. doi: 10.1038/ng0298-164
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Broglio, L., Tentorio, M., Cotelli, M.S., Mancuso, M., Vielmi, V., Gregorelli, V., et al. (2010). Белковые заболевания, связанные с поясно-конечностной мышечной дистрофией. Невролог 16, 340–352. дои: 10.1097/NRL.0b013e3181d35b39
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Браун, С. Б., Энглека, К. А., Веннинг, Дж., Мин Лу, М., и Эпштейн, Дж. А. (2005). Идентификация энхансерного элемента гипаксиальных сомитов, регулирующего экспрессию Pax3 в мигрирующих миобластах, и характеристика трансгенных мышей Cre гипаксиальных мышц. Бытие 41, 202–209. doi: 10.1002/gene.20116
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Берк, А.С., и Новицкий, Дж.Л. (2003). Новый взгляд на домены формирования паттерна в мезодерме позвоночных. Дев. Ячейка 4, 159–165. doi: 10.1016/S1534-5807(03)00033-9
doi: 10.1016/S1534-5807(03)00033-9
Полнотекстовая перекрестная ссылка | Академия Google
Камачо Ванегас, О., Bertini, E., Zhang, R.Z., Petrini, S., Minosse, C., Sabatelli, P., et al. (2001). Причиной склероатонической мышечной дистрофии Ульриха являются рецессивные мутации в коллагене VI типа. Проц. Натл. акад. науч. США 98, 7516–7521. doi: 10.1073/pnas.121027598
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Castets, P., Bertrand, A.T., Beuvin, M., Ferry, A., Le Grand, F., Castets, M., et al. (2011). Потеря сателлитных клеток и нарушение регенерации мышц при дефиците селенопротеина N. Гул. Мол. Жене. 20, 694–704. DOI: 10.1093/hmg/ddq515
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Chen, S.C., Frett, E., Marx, J., Bosnakovski, D., Reed, X., Kyba, M., et al. (2011). Снижение кинетики пролиферации миобластов мыши, сверхэкспрессирующих FRG1. PLoS ONE 6:e19780. doi: 10.1371/journal.pone.0019780
doi: 10.1371/journal.pone.0019780
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Christ, B., и Ordahl, C.P. (1995).Ранние стадии развития куриных сомитов. Анат. Эмбриол. 191, 381–396. дои: 10.1007/BF00304424
Полнотекстовая перекрестная ссылка | Академия Google
Кристиансен, С.П., и Маклун, Л.К. (2006). Влияние резекции на активность сателлитных клеток экстраокулярной мышцы кролика. Инвест. Офтальмол. Вис. науч. 47, 605–613. doi: 10.1167/iovs.05-1069
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Коллинз, К.А., Олсен, И., Заммит, П.С., Хеслоп Л., Петри А., Партридж Т.А. и соавт. (2005). Функция стволовых клеток, самообновление и поведенческая гетерогенность клеток из ниши сателлитных клеток взрослых мышц. Сотовый 122, 289–301. doi: 10.1016/j.cell.2005.05.010
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Купер, Р. Н., Тайбахш, С., Моули, В., Коссу, Г., Букингем, М., и Батлер-Браун, Г. С. (1999). Активация сателлитных клеток in vivo через Myf5 и MyoD в регенерирующих скелетных мышцах мыши. J. Cell Sci. 112 (часть 17), 2895–2901.
Н., Тайбахш, С., Моули, В., Коссу, Г., Букингем, М., и Батлер-Браун, Г. С. (1999). Активация сателлитных клеток in vivo через Myf5 и MyoD в регенерирующих скелетных мышцах мыши. J. Cell Sci. 112 (часть 17), 2895–2901.
Академия Google
Корнелисон, Д. Д., Филла, М. С., Стэнли, Х. М., Рапрегер, А. С., и Олвин, Б. Б. (2001). Синдекан-3 и синдекан-4 специфически маркируют сателлитные клетки скелетных мышц и участвуют в поддержании сателлитных клеток и регенерации мышц. Дев. биол. 239, 79–94. doi: 10.1006/dbio.2001.0416
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Корнелисон, Д.Д. и Уолд, Б. Дж. (1997). Одноклеточный анализ экспрессии регуляторных генов в покоящихся и активированных сателлитных клетках скелетных мышц мыши. Дев. биол. 191, 270–283. doi: 10.1006/dbio.1997.8721
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Couly, G.F., Coltey, P.M., and Le Douarin, N. M. (1992). Судьба развития головной мезодермы у химер перепелов и цыплят. Разработка 114, 1–15.
M. (1992). Судьба развития головной мезодермы у химер перепелов и цыплят. Разработка 114, 1–15.
Реферат PubMed | Академия Google
Чайковский, М.Т., Рассек К., Ленхард Д.К., Броль Д. и Бирчмайер К. (2014). Различная и консервативная роль передачи сигналов Dll1 в развитии черепно-лицевой и туловищной мышц. Дев. биол. 395, 307–316. doi: 10.1016/j.ydbio.2014.09.005
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Дэй, Дж. В., и Ранум, Л. П. (2005). Генетика и молекулярный патогенез миотонических дистрофий. Курс. Нейрол. Неврологи. 5, 55–59. doi: 10.1007/s11910-005-0024-1
Полнотекстовая перекрестная ссылка | Академия Google
Дэй, К., Шефер Г., Ричардсон Дж. Б., Ениколопов Г. и Яблонка-Реувени З. (2007). Экспрессия репортера Nestin-GFP определяет состояние покоя сателлитных клеток скелетных мышц. Дев. биол. 304, 246–259. doi: 10.1016/j.ydbio.2006.12.026
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Декари С. , Хамида С. Б., Мули В., Барбет Дж. П., Хентати Ф. и Батлер-Браун Г. С. (2000). Более короткие теломеры в дистрофических мышцах соответствуют обширной регенерации у маленьких детей. Нервно-мышечная. Беспорядок. 10, 113–120. дои: 10.1016/S0960-8966(99)00093-0
, Хамида С. Б., Мули В., Барбет Дж. П., Хентати Ф. и Батлер-Браун Г. С. (2000). Более короткие теломеры в дистрофических мышцах соответствуют обширной регенерации у маленьких детей. Нервно-мышечная. Беспорядок. 10, 113–120. дои: 10.1016/S0960-8966(99)00093-0
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Диксит, М., Ансо, Э., Тассин, А., Винокур, С., Ши, Р., Цянь, Х., и др. (2007). DUX4, ген-кандидат лицелопаточно-плечевой мышечной дистрофии, кодирует транскрипционный активатор PITX1. Проц. Натл. акад. науч. США 104, 18157–18162. doi: 10.1073/pnas.0708659104
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Дунхуэй, С., Shicai, C., Wei, W., Fei, L., Jianjun, J., Gang, C., et al. (2010). Функциональная модуляция сателлитных клеток в длительно денервированной мышце гортани человека. Ларингоскоп 120, 353–358. doi: 10.1002/lary.20796
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Доннер М. В., Босма Дж.Ф. и Робертсон Д.Л. (1985). Анатомия и физиология глотки. Гастроинтест. Радиол. 10, 196–212. дои: 10.1007/BF01893103
В., Босма Дж.Ф. и Робертсон Д.Л. (1985). Анатомия и физиология глотки. Гастроинтест. Радиол. 10, 196–212. дои: 10.1007/BF01893103
Полнотекстовая перекрестная ссылка | Академия Google
Дадди, В., Duguez, S., Johnston, H., Cohen, T.V., Phadke, A., Gordish-Dressman, H., et al. (2015). Мышечная дистрофия у мышей mdx представляет собой тяжелую миопатию, сочетающуюся с гипотрофией, гипертрофией и гиперплазией. Скелет. Мышца 5, 16. doi: 10.1186/s13395-015-0041-y
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Дурбидж, М., и Кэмпбелл, К.П. (2002). Мышечные дистрофии с участием комплекса дистрофин-гликопротеин: обзор современных моделей мышей. Курс.мнение Жене. Дев. 12, 349–361. doi: 10.1016/S0959-437X(02)00309-X
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Экберг О., Экман М., Эрикссон Л. И., Мальм Р., Сандман Э. и Арнер А. (2009). Модель in vitro для изучения нервно-мышечной передачи в глотке мыши. Дисфагия 24, 32–39. doi: 10.1007/s00455-008-9168-x
Дисфагия 24, 32–39. doi: 10.1007/s00455-008-9168-x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Эртекин С., Айдогду И.(2003). Нейрофизиология глотания. клин. Нейрофизиол. 114, 2226–2244. doi: 10.1016/S1388-2457(03)00237-2
Полнотекстовая перекрестная ссылка | Академия Google
Фавро, К., Иге, Д., Курвален, Дж. К., и Буэндиа, Б. (2004). Экспрессия мутантного ламина А, который вызывает мышечную дистрофию Эмери-Дрейфуса, ингибирует дифференцировку миобластов C2C12 in vitro. Мол. Клетка. биол. 24, 1481–1492. doi: 10.1128/MCB.24.4.1481-1492.2004
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Фини, С.J., McGrath, M.J., Sriratana, A., Gehrig, S.M., Lynch, G.S., D’Arcy, C.E., et al. (2015). FHL1 уменьшает дистрофию у трансгенных мышей со сверхэкспрессией гена 1 области мышечной дистрофии FSHD (FRG1). PLoS ONE 10:e0117665. doi: 10.1371/journal.pone.0117665
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Формикола, Л. , Марацци, Г., и Сассун, Д. А. (2014). Ниша экстраокулярных мышечных стволовых клеток устойчива к старению и болезням. Фронт. Стареющие нейроски. 6:328. doi: 10.3389/fnagi.2014.00328
, Марацци, Г., и Сассун, Д. А. (2014). Ниша экстраокулярных мышечных стволовых клеток устойчива к старению и болезням. Фронт. Стареющие нейроски. 6:328. doi: 10.3389/fnagi.2014.00328
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Фрок Р.Л., Кудлоу Б.А., Эванс А.М., Джеймсон С.А., Хаушка С.Д. и Кеннеди Б.К. (2006). Ламин A/C и эмерин имеют решающее значение для дифференцировки сателлитных клеток скелетных мышц. Гены Дев. 20, 486–500. doi: 10.1101/gad.1364906
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Фрай, К.С., Ли, Дж.Д., Мула, Дж., Кирби, Т.J., Jackson, J.R., Liu, F., et al. (2015). Индуцируемое истощение сателлитных клеток у взрослых малоподвижных мышей ухудшает регенеративную способность мышц, не влияя на саркопению. Нац. Мед. 21, 76–80. doi: 10.1038/nm.3710
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Фукада С., Уезуми А., Икемото М. , Масуда С., Сегава М., Танимура Н. и др. (2007). Молекулярная подпись покоящихся сателлитных клеток в скелетных мышцах взрослых. Стволовые клетки 25, 2448–2459.doi: 10.1634/стволовые клетки.2007-0019
, Масуда С., Сегава М., Танимура Н. и др. (2007). Молекулярная подпись покоящихся сателлитных клеток в скелетных мышцах взрослых. Стволовые клетки 25, 2448–2459.doi: 10.1634/стволовые клетки.2007-0019
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Фукада С., Ямагути М., Кокубо Х., Огава Р., Уэдзуми А., Йонеда Т. и другие. (2011). Hesr1 и Hesr3 необходимы для образования недифференцированных покоящихся сателлитных клеток и для поддержания числа сателлитных клеток. Разработка 138, 4609–4619. doi: 10.1242/dev.067165
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ферлинг, Д., Койффье, Л., Mouly, V., Barbet, J.P., St Guily, J.L., Taneja, K., et al. (2001). Дефектные сателлитные клетки при врожденной миотонической дистрофии. Гул. Мол. Жене. 10, 2079–2087. doi: 10.1093/hmg/19.10.2079
Полнотекстовая перекрестная ссылка | Академия Google
Габеллини, Д., Грин, М. Р. , и Таплер, Р. (2002). Несоответствующая активация гена при ЛЛПД: репрессорный комплекс связывает хромосомный повтор, удаленный в дистрофической мышце. Сотовый 110, 339–348. doi: 10.1016/S0092-8674(02)00826-7
, и Таплер, Р. (2002). Несоответствующая активация гена при ЛЛПД: репрессорный комплекс связывает хромосомный повтор, удаленный в дистрофической мышце. Сотовый 110, 339–348. doi: 10.1016/S0092-8674(02)00826-7
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Гаттаццо, Ф., Molon, S., Morbidoni, V., Braghetta, P., Blaauw, B., Urciuolo, A., et al. (2014). Циклоспорин А способствует in vivo миогенному ответу у мышей с дефицитом коллагена VI и миопатией. Фронт. Стареющие нейроски. 6:244. doi: 10.3389/fnagi.2014.00244
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Goding, G.S. Jr., Al-Sharif, K.I., and McLoon, L.K. (2005). Миоядерное дополнение к неповрежденным миофибриллам гортани у взрослых кроликов. Энн. Отол. Ринол. Ларингол. 114, 552–557. дои: 10.1177/000348940511400711
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Gustafsson, M.K., Pan, H. , Pinney, D.F., Liu, Y., Lewandowski, A., Epstein, D.J., et al. (2002). Myf5 является прямой мишенью дальнодействующей передачи сигналов Shh и регуляции Gli для спецификации мышц. Гены Дев. 16, 114–126. doi: 10.1101/gad.940702
, Pinney, D.F., Liu, Y., Lewandowski, A., Epstein, D.J., et al. (2002). Myf5 является прямой мишенью дальнодействующей передачи сигналов Shh и регуляции Gli для спецификации мышц. Гены Дев. 16, 114–126. doi: 10.1101/gad.940702
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Хак, Р. У., Шпеер, М.С., Чу, М.Л., и Тандан, Р. (1999). Поражение дыхательных мышц при миопатии Бетлема. Неврология 52, 174–176. doi: 10.1212/WNL.52.1.174
Полнотекстовая перекрестная ссылка | Академия Google
Хеберт, С.Л., Дэниел, М.Л., и Маклун, Л.К. (2013). Роль Pitx2 в поддержании фенотипа миогенных клеток-предшественников в экстраокулярных мышцах. PLoS ONE 8:e58405. doi: 10.1371/journal.pone.0058405
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Хигучи И., Каваи Х., Кавадзири М., Фукунага Х., Хорикири Т., Ниияма Т. и др. (1999). Статистически значимые различия в количестве CD24-положительных мышечных волокон и сателлитных клеток между пациентами с саркогликанопатией и соответствующими по возрасту пациентами с мышечной дистрофией Беккера. Интерн. Мед. 38, 412–415. doi: 10.2169/internalmedicine.38.412
Интерн. Мед. 38, 412–415. doi: 10.2169/internalmedicine.38.412
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Himmelreich, HA (1973). Небно-глоточная мышца млекопитающих. Гегенбаурс Морфол.Ярб. 119, 172–212.
Академия Google
Хоффман, Е. П., Браун, Р. Х. мл., и Кункель, Л. М. (1987). Дистрофин: белковый продукт очага мышечной дистрофии Дюшенна. Сотовый 51, 919–928. дои: 10.1016/0092-8674(87)
-4Полнотекстовая перекрестная ссылка | Академия Google
Икемото М., Фукада С., Уезуми А., Масуда С., Миёси Х., Ямамото Х. и др. (2007). Аутологичная трансплантация сателлитных клеток SM/C-2.6(+), трансдуцированных кДНК микродистрофина CS1 с помощью лентивирусного вектора, мышам mdx. Мол. тер. 15, 2178–2185. doi: 10.1038/sj.mt.6300295
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ипполито Дж., Арпке Р.В., Хайдер К.Т., Чжан Дж. и Киба М. (2012). Неоднородность сателлитных клеток, обнаруженная с помощью G-Tool, открытого алгоритма для количественной оценки миогенеза с помощью анализов образования колоний. Скелет. Мышца 2, 13. doi: 10.1186/2044-5040-2-13
(2012). Неоднородность сателлитных клеток, обнаруженная с помощью G-Tool, открытого алгоритма для количественной оценки миогенеза с помощью анализов образования колоний. Скелет. Мышца 2, 13. doi: 10.1186/2044-5040-2-13
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Янссен, И., Heymsfield, S.B., Wang, Z.M., and Ross, R. (2000). Масса и распределение скелетных мышц у 468 мужчин и женщин в возрасте 18-88 лет. Дж. Заявл. Физиол. 89, 81–88.
Реферат PubMed | Академия Google
Цзян, К., Вен, Ю., Курода, К., Хэннон, К., Рудницкий, М.А., и Куанг, С. (2014). Дефицит передачи сигналов Notch лежит в основе возрастного истощения сателлитных клеток при мышечной дистрофии. Дис. Модель мех. 7, 997–1004. doi: 10.1242/dmm.015917
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Джо, А.В., Йи, Л., Натараджан, А., Ле Гранд, Ф., Со, Л., Ван, Дж., и соавт. (2010). Повреждение мышц активирует резидентные фибро/адипогенные предшественники, которые облегчают миогенез. Нац. Клеточная биол. 12, 153–163. дои: 10.1038/ncb2015
Нац. Клеточная биол. 12, 153–163. дои: 10.1038/ncb2015
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Джоуан, Л., Рошфорд, Д., Шуто, А., Карни, Э., Дэвид, К., Дион, П.А., и соавт. (2014). 18-аланиновый повтор при тяжелой форме окулофарингеальной мышечной дистрофии. Кан. Дж. Нейрол. науч. 41, 508–511. дои: 10.1017/S0317167100018588
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кафадар К.А., Йи Л., Ахмад Ю., Со Л., Росси Ф. и Павлат Г.К. (2009). Экспрессия Sca-1 необходима для эффективного ремоделирования внеклеточного матрикса во время регенерации скелетных мышц. Дев. биол. 326, 47–59. doi: 10.1016/j.ydbio.2008.10.036
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Каллестад, К.M., Hebert, S.L., McDonald, A.A., Daniel, M.L., Cu, S.R., и McLoon, L.K. (2011). Сохранение экстраокулярных мышц при старении и мышечных дистрофиях: гипотеза миогенных клеток-предшественников. Экспл. Сотовый рез. 317, 873–885. doi: 10.1016/j.yexcr.2011.01.018
Экспл. Сотовый рез. 317, 873–885. doi: 10.1016/j.yexcr.2011.01.018
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Kaminski, H.J., al-Hakim, M., Leigh, R.J., Katirji, M.B., и Ruff, R.L. (1992). Экстраокулярные мышцы не сохраняются при дистрофии Дюшенна на поздних стадиях. Энн.Нейрол. 32, 586–588. doi: 10.1002/ana.410320418
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Каваи М., Сайтсу К., Ямасита Х. и Мията Х. (2012). Возрастные изменения пролиферации сателлитных клеток путем компенсаторной активации мышц диафрагмы крыс. Биомед. Рез. 33, 167–173. doi: 10.2220/biomedres.33.167
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Киф, А.С., Лоусон, Дж.А., Флайгар, С.Д., Fox, Z.D., Colasanto, M.P., Mathew, S.J., et al. (2015). Мышечные стволовые клетки вносят вклад в миофибриллы у сидячих взрослых мышей. Нац. коммун. 6, 7087. doi: 10. 1038/ncomms8087
1038/ncomms8087
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Келли, Р. Г., Джером-Маевска, Л. А., и Папайоанноу, В. Е. (2004). Ген-кандидат del22q11.2 Tbx1 регулирует бранхиомерный миогенез. Гул. Мол. Жене. 13, 2829–2840. doi: 10.1093/hmg/ddh404
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ковальёв, В., Marcowycz, A., Ansseau, E., Conde, C.B., Sauvage, S., Matteotti, C., et al. (2007). Ген DUX4 в локусе FSHD1A кодирует проапоптотический белок. Нервно-мышечная. Беспорядок. 17, 611–623. doi: 10.1016/j.nmd.2007.04.002
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Куанг С., Курода К., Ле Гранд Ф. и Рудницки М. А. (2007). Асимметричное самообновление и коммитирование сателлитных стволовых клеток в мышцах. Сотовый 129, 999–1010. doi: 10.1016/j.моб.2007.03.044
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кудряшова Э. , Крамерова И. и Спенсер М. Дж. (2012). Старение сателлитных клеток лежит в основе миопатии на мышиной модели поясно-конечностной мышечной дистрофии 2Н. Дж. Клин. Вкладывать деньги. 122, 1764–1776 гг. DOI: 10.1172/JCI59581
, Крамерова И. и Спенсер М. Дж. (2012). Старение сателлитных клеток лежит в основе миопатии на мышиной модели поясно-конечностной мышечной дистрофии 2Н. Дж. Клин. Вкладывать деньги. 122, 1764–1776 гг. DOI: 10.1172/JCI59581
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кюн, У., Гюндель, М., Кнот, А., Кервитц, Ю., Рюдель, С., и Вале, Э. (2009). Длина поли(А)-хвоста контролируется ядерным поли(А)-связывающим белком, регулирующим взаимодействие между поли(А)-полимеразой и фактором специфичности расщепления и полиаденилирования. Дж. Биол. хим. 284, 22803–22814. doi: 10.1074/jbc.M109.018226
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кумай Ю., Ито Т., Миямару С. и Юмото Э. (2007). Модуляция MyoD- и Ki-67-позитивных сателлитных клеток в кратковременно денервированной тирочерпаловидной мышце крысы. Ларингоскоп 117, 2063–2067. дои: 10.1097/MLG.0b013e318133a13c
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
ЛаФрамбуаз, В. A., Guthrie, R.D., Scalise, D., Elborne, V., Bombach, K.L., Armanious, C.S., et al. (2003). Влияние мышечного происхождения и фенотипа на экспрессию генов, специфичных для мышц, сателлитных клеток. Дж. Мол. Клетка. Кардиол. 35, 1307–1318. doi: 10.1016/S0022-2828(03)00245-1
A., Guthrie, R.D., Scalise, D., Elborne, V., Bombach, K.L., Armanious, C.S., et al. (2003). Влияние мышечного происхождения и фенотипа на экспрессию генов, специфичных для мышц, сателлитных клеток. Дж. Мол. Клетка. Кардиол. 35, 1307–1318. doi: 10.1016/S0022-2828(03)00245-1
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ли, К., Клецин, Х., Коннор, Н.П., Шульц, Э., Чемберлен, К.С., и Блесс, Д.М. (2012). Влияние старения на регенерацию щиточерпаловидных мышц. Ларингоскоп 122, 2800–2807. doi: 10.1002/лари.23589
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Леппер, К., Партридж, Т.А., и Фан, К.М. (2011). Абсолютная потребность в Pax7-позитивных сателлитных клетках для регенерации скелетных мышц, вызванной острой травмой. Разработка 138, 3639–3646. doi: 10.1242/dev.067595
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Лескодрон, Л., Пелтекян, Э. , Фонтен-Перю, Дж., Paulin, D., Zampieri, M., Garcia, L., et al. (1999). Макрофаги, переносимые кровью, необходимы для запуска регенерации мышц после трансплантации мышц. Нервно-мышечная. Беспорядок. 9, 72–80. doi: 10.1016/S0960-8966(98)00111-4
, Фонтен-Перю, Дж., Paulin, D., Zampieri, M., Garcia, L., et al. (1999). Макрофаги, переносимые кровью, необходимы для запуска регенерации мышц после трансплантации мышц. Нервно-мышечная. Беспорядок. 9, 72–80. doi: 10.1016/S0960-8966(98)00111-4
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Л’Оноре, А., Уиметт, Дж. Ф., Лавертю-Джолин, М., и Друэн, Дж. (2010). Pitx2 определяет альтернативные пути, действующие через MyoD во время миогенеза конечностей и сомитов. Разработка 137, 3847–3856.
Реферат PubMed | Академия Google
Макинтош, Б. Р., Гардинер, П. Ф., и МакКомас, А. Дж. (2006). Скелетные мышцы: форма и функция . Шампейн, Иллинойс: Кинетика человека.
Академия Google
Малатеста, М., Джаньяково, М., Ренна, Л.В., Кардани, Р., Меола, Г., и Пелличчари, К. (2011). Культивируемые миобласты пациентов, страдающих миотонической дистрофией 2 типа, демонстрируют признаки, связанные со старением: ультраструктурные признаки. евро. Дж. Гистохим. 55, е26. doi: 10.4081/ejh.2011.e26
евро. Дж. Гистохим. 55, е26. doi: 10.4081/ejh.2011.e26
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Мальмгрен, Л. Т., Фишер, П. Дж., Джонс, К. Э., Букман, Л. М., и Уно, Т. (2000). Численная плотность миоядер и сателлитных клеток в типах мышечных волокон в стареющей тиреочерпаловидной мышце человека: иммуногистохимическое и стереологическое исследование с использованием конфокальной лазерной сканирующей микроскопии. Отоларингол. Хирургия головы и шеи. 123, 377–384. doi: 10.1067/мн.2000.109487
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Маркес, М.Дж., Ферретти Р., Вомеро В.У., Минател Э. и Нето Х.С. (2007). Внутренние мышцы гортани избавлены от мионекроза в модели мышечной дистрофии Дюшенна у мышей mdx. Мышечный нерв 35, 349–353. doi: 10.1002/mus.20697
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Martin, B.J., Corlew, M.M., Wood, H. , Olson, D., Golopol, L.A., Wingo, M., et al. (1994). Связь нарушения глотания и аспирационной пневмонии. Дисфагия 9, 1–6.дои: 10.1007/BF00262751
, Olson, D., Golopol, L.A., Wingo, M., et al. (1994). Связь нарушения глотания и аспирационной пневмонии. Дисфагия 9, 1–6.дои: 10.1007/BF00262751
Полнотекстовая перекрестная ссылка | Академия Google
Мастрояннопулос, Н.П., Шаммас, К., и Филакту, Л.А. (2010). Рассмотрение патогенеза задержки ядра РНК при миотонической дистрофии. Биол. Мобильный 102, 515–523. дои: 10.1042/BC20100040
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Mathew, S.J., Hansen, J.M., Merrell, A.J., Murphy, M.M., Lawson, J.A., Hutcheson, D.A., et al. (2011). Фибробласты соединительной ткани и Tcf4 регулируют миогенез. Разработка 138, 371–384. дои: 10.1242/dev.057463
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Мауро, А. (1961). Сателлитная клетка скелетных мышечных волокон. Дж. Биофиз. Биохим. Цитол. 9, 493–495. doi: 10.1083/jcb.9.2.493
Полнотекстовая перекрестная ссылка | Академия Google
McCarthy, J. J., Mula, J., Miyazaki, M., Erfani, R., Garrison, K., Farooqui, A.B., et al. (2011). Эффективная гипертрофия волокон в скелетных мышцах с истощением сателлитных клеток. Разработка 138, 3657–3666. doi: 10.1242/dev.068858
J., Mula, J., Miyazaki, M., Erfani, R., Garrison, K., Farooqui, A.B., et al. (2011). Эффективная гипертрофия волокон в скелетных мышцах с истощением сателлитных клеток. Разработка 138, 3657–3666. doi: 10.1242/dev.068858
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Макдермотт, А., Густафссон, М., Элсам, Т., Хуэй, К.С., Эмерсон, С.П. мл., и Борицкий, А.Г. (2005). Gli2 и Gli3 выполняют избыточную и контекстно-зависимую функцию в формировании скелетных мышц. Разработка 132, 345–357. doi: 10.1242/dev.01537
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Макдональдс, А.А., Кунц, М.Д., и Маклун, Л.К. (2014). Дистрофические изменения экстраокулярных мышц после гамма-облучения у мышей mdx:utrophin(+/-). PLoS ONE 9:e86424. doi: 10.1371/journal.pone.0086424
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
МакГичи, Дж., и Оллбрук, Д. (1978). Пролиферация клеток в скелетных мышцах после денервации или тенотомии. Серия авторадиографических исследований. Рез. клеточной ткани. 193, 259–267. дои: 10.1007/BF00209039
Серия авторадиографических исследований. Рез. клеточной ткани. 193, 259–267. дои: 10.1007/BF00209039
Полнотекстовая перекрестная ссылка | Академия Google
Макленнан, И.С. (1996). Дегенерирующие и регенерирующие скелетные мышцы содержат несколько субпопуляций макрофагов с различным пространственным и временным распределением. Дж. Анат. 188 (ч. 1), 17–28.
Реферат PubMed | Академия Google
МакЛун, Л.К., Торстенсон, К.М., Соломон, А., и Льюис, М.П. (2007). Миогенные клетки-предшественники в черепно-лицевых мышцах. Оральный дис. 13, 134–140.
Академия Google
McLoon, L.K., and Wirtschafter, J. (2002a).Активированные сателлитные клетки присутствуют в неповрежденных экстраокулярных мышцах взрослых мышей. Пер. Являюсь. Офтальмол. соц. 100, 119–123; обсуждение 123–114.
Реферат PubMed | Академия Google
McLoon, L.K., и Wirtschafter, JD (2002b). Непрерывное миоядерное добавление к одиночным экстраокулярным миофибриллам у неповрежденных взрослых кроликов. Мышечный нерв 25, 348–358. doi: 10.1002/mus.10056
Мышечный нерв 25, 348–358. doi: 10.1002/mus.10056
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
МакЛун, Л.К. и Виртшафтер Дж. (2003). Активированные сателлитные клетки в экстраокулярных мышцах нормальных взрослых обезьян и человека. Инвест. Офтальмол. Вис. науч. 44, 1927–1932 гг. doi: 10.1167/iovs.02-0673
Полнотекстовая перекрестная ссылка | Академия Google
Мейнке, П., Шнайдерат, П., Срсен, В., Корфали, Н., Ле Тхань, П., Коуэн, Г.Дж., и соавт. (2015). Аномальная пролиферация и спонтанная дифференцировка миобластов у женщины-носительницы Х-сцепленной мышечной дистрофии Эмери-Дрейфуса с симптомами. Нервно-мышечная. Беспорядок. 25, 127–136.
Академия Google
Меррелл, А. Дж., Эллис, Б. Дж., Фокс, З. Д., Лоусон, Дж. А., Вайс, Дж. А., и Кардон, Г. (2015). Мышечная соединительная ткань контролирует развитие диафрагмы и является источником врожденных диафрагмальных грыж. Нац. Жене. 47, 496–504. doi: 10.1038/ng.3250
47, 496–504. doi: 10.1038/ng.3250
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Миллер, А. Дж. (2002). Оральные и глоточные рефлексы в нервной системе млекопитающих: их разнообразие по сложности и ключевая роль языка. Крит. Преподобный Орал Биол. Мед. 13, 409–425. дои: 10.1177/154411130201300505
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Minetti, G.C., Colussi, C., Adami, R., Serra, C., Mozzetta, C., Parente, V., et al. (2006). Функциональное и морфологическое восстановление дистрофических мышц у мышей, получавших ингибиторы деацетилазы. Нац. Мед. 12, 1147–1150. doi: 10.1038/nm1479
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Миттельбронн, М., Салливан, Т., Стюарт, К.Л., и Борнеманн, А. (2008). Мионуклеарная дегенерация у мышей с нулевым LMNA. Патология головного мозга. 18, 338–343. doi: 10.1111/j.1750-3639.2008.00123. x
x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Миягоэ-Сузуки Ю., Масубучи Н., Миямото К., Вада М. Р., Юаса С., Сайто Ф. и др. (2009). Снижение пролиферативной активности первичных POMGnT1-нулевых миобластов in vitro. Мех. Дев. 126, 107–116. doi: 10.1016/j.mod.2008.12.001
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Мохонова Э.И., Авлиякулов Н.К., Крамерова И., Кудряшова Э., Хайкинсон М.Дж. и Спенсер М.Дж. (2015). Убиквитинлигаза Е3 TRIM32 регулирует пролиферацию миобластов, контролируя оборот NDRG2. Гул. Мол. Жене. 24, 2873–2883. DOI: 10.1093/hmg/ddv049
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Монако, А. П., Бертельсон, С. Дж., Лихти-Галлати, С., Мозер, Х., и Кункель, Л.М. (1988). Объяснение фенотипических различий между пациентами с частичными делециями локуса DMD. Геномика 2, 90–95. дои: 10.1016/0888-7543(88)-9
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Мутоосами, Р. К., и Дитрих, С. (2002). Различные регуляторные каскады для миогенеза головы и туловища. Разработка 129, 573–583.
К., и Дитрих, С. (2002). Различные регуляторные каскады для миогенеза головы и туловища. Разработка 129, 573–583.
Академия Google
Морган, Дж. Э., и Заммит, П.С. (2010). Прямое влияние патогенной мутации на функцию сателлитных клеток при мышечной дистрофии. Экспл. Сотовый рез. 316, 3100–3108. doi: 10.1016/j.yexcr.2010.05.014
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Мотохаси, Н., и Асакура, А. (2014). Гетерогенность и самообновление мышечных сателлитных клеток. Фронт. Сотовый Дев. биол. 2:1. doi: 10.3389/fcell.2014.00001
Полнотекстовая перекрестная ссылка | Академия Google
Мули, В., Aamiri, A., Bigot, A., Cooper, R.N., Donna, S. Di Furling, D., et al. (2005). Митотические часы в регенерации скелетных мышц, заболеваниях и генной терапии, опосредованной клетками. Acta Physiol. Сканд. 184, 3–15. doi: 10.1111/j.1365-201X.2005.01417.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Мюнстерберг А. Е., Китаевский Дж., Бамкрот Д.А., МакМахон А.П. и Лассар А.Б. (1995). Комбинаторная передача сигналов Sonic hedgehog и членами семейства Wnt индуцирует экспрессию миогенного гена bHLH в сомитах. Гены Дев. 9, 2911–2922. doi: 10.1101/gad.9.23.2911
Е., Китаевский Дж., Бамкрот Д.А., МакМахон А.П. и Лассар А.Б. (1995). Комбинаторная передача сигналов Sonic hedgehog и членами семейства Wnt индуцирует экспрессию миогенного гена bHLH в сомитах. Гены Дев. 9, 2911–2922. doi: 10.1101/gad.9.23.2911
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Мерфи, М.М., Лоусон, Дж.А., Мэтью, С.Дж., Хатчесон, Д.А., и Кардон, Г. (2011). Сателлитные клетки, фибробласты соединительной ткани и их взаимодействие имеют решающее значение для регенерации мышц. Разработка 138, 3625–3637. doi: 10.1242/dev.064162
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Натан, Э., Монович А., Тирош-Финкель Л., Харрельсон З., Руссо Т., Ринон А. и соавт. (2008). Вклад клеток мезодермы, экспрессирующих Islet1, в отдельные бранхиомерные мышцы обнаруживает значительную гетерогенность в развитии мышц головы. Разработка 135, 647–657. doi: 10.1242/dev.007989
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Нигро Г. , Коми Л.И., Политано Л. и Бейн Р.Дж. (1990). Заболеваемость и эволюция кардиомиопатии при мышечной дистрофии Дюшенна. Междунар. Дж. Кардиол. 26, 271–277. дои: 10.1016/0167-5273(90)
, Коми Л.И., Политано Л. и Бейн Р.Дж. (1990). Заболеваемость и эволюция кардиомиопатии при мышечной дистрофии Дюшенна. Междунар. Дж. Кардиол. 26, 271–277. дои: 10.1016/0167-5273(90)
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ноден Д.М., Маркучо Р., Борицкий А.Г. и Эмерсон С.П. мл. (1999). Дифференциация черепно-лицевых мышц птиц: I. Паттерны ранней экспрессии регуляторных генов и синтеза тяжелых цепей миозина. Дев. Дин. 216, 96–112.
Реферат PubMed | Академия Google
Олсон, Э. Н., и Кляйн, У. Х. (1994).Факторы bHLH в развитии мышц: крайние сроки и обязательства, что оставить, а что исключить. Гены Дев. 8, 1–8. doi: 10.1101/gad.8.1.1
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Оно, Ю., Болдрин, Л., Кнопп, П., Морган, Дж. Э., и Заммит, П. С. (2010). Мышечные сателлитные клетки представляют собой функционально гетерогенную популяцию как в сомитных, так и в бранхиомерных мышцах. Дев. биол. 337, 29–41. doi: 10.1016/j.ydbio.2009.10.005
Дев. биол. 337, 29–41. doi: 10.1016/j.ydbio.2009.10.005
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Pacheco-Pinedo, E.C., Budak, M.T., Zeiger, U., Jørgensen, L.H., Bogdanovich, S., Schrøder, H.D., et al. (2009). Транскрипционные и функциональные различия в популяциях стволовых клеток, выделенных из экстраокулярных мышц и мышц конечностей. Физиол. Геномика 37, 35–42. doi: 10.1152/physiolgenomics.00051.2008
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Пако, С., Феррер, И., Jou, C., Cusi, V., Corbera, J., Torner, F., et al. (2012). Атрофия и регенерация мышечных волокон сосуществуют в мышцах человека с дефицитом коллагена VI: роль передачи сигналов кальпаина-3 и ядерного фактора-каппаВ. Дж. Невропатол. Эксп. Нейрол. 71, 894–906. дои: 10.1097/NEN.0b013e31826c6f7b
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Паллафакчина, Г., Блаау, Б. , и Скьяффино, С. (2013). Роль сателлитных клеток в росте мышц и поддержании мышечной массы. Нутр. Метаб. Кардиовас. Дис. 23(Прил. 1), С12–С18. doi: 10.1016/j.numecd.2012.02.002
, и Скьяффино, С. (2013). Роль сателлитных клеток в росте мышц и поддержании мышечной массы. Нутр. Метаб. Кардиовас. Дис. 23(Прил. 1), С12–С18. doi: 10.1016/j.numecd.2012.02.002
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Pavlath, G.K., Thaloor, D., Rando, T.A., Cheong, M., English, A.W., and Zheng, B. (1998). Гетерогенность клеток-предшественников мышц во взрослых скелетных мышцах с различной регенеративной способностью. Дев. Дин. 212, 495–508. doi: 10.1002/(SICI)1097-0177(199808)212:4<495::AID-AJA3>3.3.CO;2-I
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Пери, С., Mamchaoui, K., Mouly, V., Blot, S., Bouazza, B., Thornell, L.-E., et al. (2006). Преждевременная пролиферативная остановка перстнеглоточных миобластов при окулофарингеальной мышечной дистрофии: терапевтические перспективы трансплантации аутологичных миобластов. Нервно-мышечная. Беспорядок. 16, 770–781. doi: 10. 1016/j.nmd.2006.07.022
1016/j.nmd.2006.07.022
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Пери, С., Тролле, К., Мули, В., Ванно, В., Мамчауи, К., Буацца, Б., и другие. (2014).Трансплантация аутологичных миобластов при окулофарингеальной мышечной дистрофии: клиническое исследование фазы I/IIa. Мол. тер. 22, 219–225. doi: 10.1038/mt.2013.155
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Петроф, Б.Дж., Шрагер, Дж.Б., Стедман, Х.Х., Келли, А.М., и Суини, Х.Л. (1993). Дистрофин защищает сарколемму от стрессов, возникающих при сокращении мышц. Проц. Натл. акад. науч. США 90, 3710–3714. doi: 10.1073/pnas.90.8.3710
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Пикеринг, М., и Джонс, Дж. Ф. (2002). Диафрагма: две физиологические мышцы в одной. Дж. Анат. 201, 305–312. doi: 10.1046/j.1469-7580.2002.00095.x
Полнотекстовая перекрестная ссылка | Академия Google
Портер, Дж. Д., и Бейкер, Р. С. (1996). Мышцы другого «цвета»: необычные свойства экстраокулярных мышц могут предрасполагать или защищать их при нейрогенных и миогенных заболеваниях. Неврология 46, 30–37. doi: 10.1212/WNL.46.1.30
Д., и Бейкер, Р. С. (1996). Мышцы другого «цвета»: необычные свойства экстраокулярных мышц могут предрасполагать или защищать их при нейрогенных и миогенных заболеваниях. Неврология 46, 30–37. doi: 10.1212/WNL.46.1.30
Полнотекстовая перекрестная ссылка | Академия Google
Портер, Дж. Д., Рафаэль, Дж. А., Рагуза, Р. Дж., Брюкнер, Дж. К., Трикетт, Дж. И., и Дэвис, К. Э. (1998). Сохранение экстраокулярных мышц при дистрофинопатии утрачивается у мышей, лишенных атрофина и дистрофина. J. Cell Sci. 111 (часть 13), 1801–1811 гг.
Реферат PubMed | Академия Google
Quijano-Roy, S., Avila-Smirnow, D., Carlier, R.Y., и группа исследования мышц Wb-Mri.(2012). Протокол МРТ мышц всего тела: распознавание образов при НМ с ранним началом. Нервно-мышечная. Беспорядок. 22 (Прил. 2), S68–S84. doi: 10.1016/j.nmd.2012.08.003
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Рай М., Нонгтомба У. и Граундс М. Д. (2014). Дегенерация и регенерация скелетных мышц у мышей и мух. Курс. Вверх. Дев. биол. 108, 247–281. doi: 10.1016/B978-0-12-3
Д. (2014). Дегенерация и регенерация скелетных мышц у мышей и мух. Курс. Вверх. Дев. биол. 108, 247–281. doi: 10.1016/B978-0-12-3
-9.00007-3
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Рэндольф, М.Э., Луо, К., Хо, Дж., Вест, К.Е., Соколофф, А.Дж., и Павлат, Г.К. (2014). Старение и мышечная дистрофия по-разному влияют на мышцы глотки мышей в зависимости от региона. J. Physiol. 592, 5301–5315. doi: 10.1113/jphysiol.2014.280420
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Рэндольф, М.Э., Филлипс, Б.Л., Чу, Х.Дж., Вест, К.Е., Вера, Ю., и Павлат, Г.К. (2015). Фарингеальные сателлитные клетки подвергаются миогенезу в базовых условиях и необходимы для поддержания глоточной мускулатуры. Стволовые клетки doi: 10.1002/stem.2098 [Epub перед печатью].
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Реле Ф., Монтаррас Д., Заффран С., Гейро-Морель Б. , Роканкур Д., Таджбахш С. и др. (2006). Pax3 и Pax7 имеют разные и перекрывающиеся функции во взрослых клетках-предшественниках мышц. J. Cell Biol. 172, 91–102. doi: 10.1083/jcb.200508044
, Роканкур Д., Таджбахш С. и др. (2006). Pax3 и Pax7 имеют разные и перекрывающиеся функции во взрослых клетках-предшественниках мышц. J. Cell Biol. 172, 91–102. doi: 10.1083/jcb.200508044
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ренна, Л.В., Кардани Р., Ботта А., Росси Г., Фоссати Б., Коста Э. и др. (2014). Преждевременное старение в первичных мышечных культурах миотонической дистрофии 2 типа не связано с индукцией p16. евро. Дж. Гистохим. 58, 2444. doi: 10.4081/ejh.2014.2444
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Робинсон, Д. О., Хамманс, С. Р., Рид, С. П., и Силлиборн, Дж. (2005). Окулофарингеальная мышечная дистрофия (OPMD): анализ последовательности экспансии гена PABPN1 у 86 пациентов выявил 13 различных типов экспансии и дополнительные доказательства неравной рекомбинации как мутационного механизма. Гул. Жене. 116, 267–271. doi: 10.1007/s00439-004-1235-2
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Рошето, П. , Гайро-Морель, Б., Сигл-Кашеденье, И., Бласко, М.А., и Таджбахш, С. (2012). Субпопуляция взрослых стволовых клеток скелетных мышц сохраняет все нити матричной ДНК после клеточного деления. Сотовый 148, 112–125. doi: 10.1016/j.cell.2011.11.049
, Гайро-Морель, Б., Сигл-Кашеденье, И., Бласко, М.А., и Таджбахш, С. (2012). Субпопуляция взрослых стволовых клеток скелетных мышц сохраняет все нити матричной ДНК после клеточного деления. Сотовый 148, 112–125. doi: 10.1016/j.cell.2011.11.049
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Роджерс, Дж.T., King, K.Y., Brett, J.O., Cromie, M.J., Charville, G.W., Maguire, K.K., et al. (2014). mTORC1 контролирует адаптивный переход покоящихся стволовых клеток из G0 в G(Alert). Природа 510, 393–396. doi: 10.1038/nature13255
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Росалес, X. К., Малик, В., Снех, А., Чен, Л., Льюис, С., Кота, Дж., и др. (2013). Нарушение регенерации при LGMD2A поддерживается повышенным содержанием PAX7-позитивных сателлитных клеток и нарушением регуляции специфических для мышц микроРНК. Мышечный нерв 47, 731–739. doi: 10.1002/mus.23669
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Розен, Г. Д., Санес, Дж.Р., ЛаШанс, Р., Каннингем, Дж.М., Роман, Дж., и Дин, Д.К. (1992). Роль интегрина VLA-4 и его контррецептора VCAM-1 в миогенезе. Сотовый 69, 1107–1119. дои: 10.1016/0092-8674(92)
Д., Санес, Дж.Р., ЛаШанс, Р., Каннингем, Дж.М., Роман, Дж., и Дин, Д.К. (1992). Роль интегрина VLA-4 и его контррецептора VCAM-1 в миогенезе. Сотовый 69, 1107–1119. дои: 10.1016/0092-8674(92)
Полнотекстовая перекрестная ссылка | Академия Google
Рубесин, С. Э., Джессурун, Дж., Робертсон, Д., Джонс, Б., Босма, Дж. Ф., и Доннер, М. В. (1987). Линии глотки. Рентгенография 7, 217–237. doi: 10.1148/радиография.7.2.3448633
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Сакко А., Муркиоти Ф., Тран Р., Чой Дж., Ллевеллин М., Крафт П. и др. (2010). Короткие теломеры и истощение стволовых клеток моделируют мышечную дистрофию Дюшенна у мышей mdx/mTR. Сотовый 143, 1059–1071. doi: 10.1016/j.cell.2010.11.039
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Солсбери, Э., Schoser, B., Schneider-Gold, C., Wang, G.L., Huichalaf, C., Jin, B., et al. (2009). Экспрессия повторов РНК CCUG нарушает регуляцию трансляции и деградации белков у пациентов с миотонической дистрофией 2. утра. Дж. Патол. 175, 748–762. doi: 10.2353/ajpath.2009.0
утра. Дж. Патол. 175, 748–762. doi: 10.2353/ajpath.2009.0
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Sambasivan, R., Gayraud-Morel, B., Dumas, G., Cimper, C., Paisant, S., Kelly, R., et al. (2009). Отдельные регуляторные каскады управляют судьбами клеток-предшественников экстраокулярных мышц и мышц глоточной дуги. Дев. Мобильный 16, 810–821. doi: 10.1016/j.devcel.2009.05.008
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Sambasivan, R., Yao, R., Kissenpfennig, A., Wittenberghe, L. Van, Paldi, A., Gayraud-Morel, B., et al. (2011). Сателлитные клетки, экспрессирующие Pax7, незаменимы для регенерации скелетных мышц взрослых. Разработка 138, 3647–3656. doi: 10.1242/dev.067587
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Сато, Т., Роканкур Д., Маркес Л., Торстейнсдоттир С. и Букингем М. (2010). Регуляторный каскад Pax3/Dmrt2/Myf5 функционирует в начале миогенеза. Генетика PLoS. 6:e1000897. doi: 10.1371/journal.pgen.1000897
Генетика PLoS. 6:e1000897. doi: 10.1371/journal.pgen.1000897
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Шмальбрух, Х. (1976). Морфология регенерации скелетных мышц крысы. Тканевая клетка 8, 673–692. дои: 10.1016/0040-8166(76)-2
Полнотекстовая перекрестная ссылка | Академия Google
Сил, П., Sabourin, L.A., Girgis-Gabardo, A., Mansouri, A., Gruss, P., and Rudnicki, M.A. (2000). Pax7 необходим для спецификации миогенных сателлитных клеток. Сотовый 102, 777–786. doi: 10.1016/S0092-8674(00)00066-0
Полнотекстовая перекрестная ссылка | Академия Google
Шервуд, Р.И., Кристенсен, Дж.Л., Конбой, И.М., Конбой, М.Дж., Рандо, Т.А., Вайсман, И.Л., и соавт. (2004). Выделение миогенных предшественников взрослых мышей: функциональная гетерогенность клеток внутри и приживляющихся скелетных мышц. Сотовый 119, 543–554. doi: 10.1016/j.cell.2004.10.021
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Шинин В. , Гайро-Морель Б., Гомес Д. и Таджбахш С. (2006). Асимметричное деление и косегрегация нитей матричной ДНК во взрослых мышечных сателлитных клетках. Нац. Клеточная биол. 8, 677–687. дои: 10.1038/ncb1425
, Гайро-Морель Б., Гомес Д. и Таджбахш С. (2006). Асимметричное деление и косегрегация нитей матричной ДНК во взрослых мышечных сателлитных клетках. Нац. Клеточная биол. 8, 677–687. дои: 10.1038/ncb1425
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Шиннерс, М. Дж., Годинг, Г. С., и Маклун, Л.К. (2006). Влияние перерезки возвратного гортанного нерва на гортанные мышцы взрослых кроликов. Отоларингол. Хирургия головы и шеи. 134, 413–418. doi: 10.1016/j.otohns.2005.11.037
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Шинонага, К., Фукуда, М., Судзуки, Ю., Хигаки, Т., Исида, Ю., Исии, Э., и другие. (2008). Оценка функции глотания при мышечной дистрофии Дюшенна. Дев. Мед. Детский Нейрол. 50, 478–480. doi: 10.1111/j.1469-8749.2008.00478.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Шулер, К.Ф., и Далримпл, К.Р. (2001). Молекулярная регуляция дифференцировки языка и черепно-лицевых мышц. Крит. Преподобный Орал Биол. Мед. 12, 3–17. дои: 10.1177/10454411010120010201
Крит. Преподобный Орал Биол. Мед. 12, 3–17. дои: 10.1177/10454411010120010201
Полнотекстовая перекрестная ссылка | Академия Google
Снайдер, Л., Генг, Л.Н., Леммерс, Р.Дж., Киба, М., Уэр, С.Б., Нельсон, А.М., и соавт. (2010). Плечево-лицевая дистрофия: неполное подавление ретротранспонированного гена. Генетика PLoS. 6:e1001181. doi: 10.1371/journal.pgen.1001181
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Стедман, Х.Х., Суини, Х.Л., Шрагер, Дж.Б., Магуайр, Х.К., Панеттьери, Р.А., Петроф, Б., и другие. (1991). Диафрагма мыши mdx воспроизводит дегенеративные изменения мышечной дистрофии Дюшенна. Природа 352, 536–539. дои: 10.1038/352536a0
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Штуэльсац, П., Кейре П., Алмули Р. и Яблонка-Реувени З. (2012). Современный атлас диафрагмы мыши: миогенность, васкуляризация и соединение Pax3. J. Histochem. Цитохим. 60, 638–657. дои: 10.1369/0022155412452417
дои: 10.1369/0022155412452417
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Stuelsatz, P., Shearer, A., Li, Y., Muir, L.A., Ieronimakis, N., Shen, Q.W., et al. (2014). Сателлитные клетки экстраокулярных мышц представляют собой высокоэффективные миодвигатели, сохраняющие эффективную регенеративную способность при дефиците дистрофина. Дев. биол. 397, 31–44. doi: 10.1016/j.ydbio.2014.08.035
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Тайбахш С., Роканкур Д., Коссу Г. и Букингем М. (1997). Переопределение генетических иерархий, контролирующих скелетный миогенез: pax-3 и Myf-5 действуют выше MyoD. Сотовый 89, 127–138.
Реферат PubMed | Академия Google
Тамаки Т., Акацука А., Андо К., Накамура Ю., Мацузава Х., Хотта Т. и др.(2002). Выявление миогенно-эндотелиальных клеток-предшественников в интерстициальных пространствах скелетных мышц. J. Cell Biol. 157, 571–577. doi: 10.1083/jcb.200112106
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Терно, Дж. П., и Порталье, П. (1993). Влияние миобластов языка на диссоциированные подъязычные мотонейроны крыс в культуре. Междунар. Дж. Дев. Неврологи. 11, 33–48. дои: 10.1016/0736-5748(93)-А
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Торнелл, Л.Э., Линдстом М., Рено В., Кляйн А., Мули В., Ансвед Т. и соавт. (2009). Дисфункция сателлитных клеток способствует прогрессирующей атрофии мышц при миотонической дистрофии 1 типа. Нейропатол. заявл. Нейробиол. 35, 603–613. doi: 10.1111/j.1365-2990.2009.01014.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Тилеман, А. А., Кнюйт, С., ван Влит, Дж., де Сварт, Б. Дж., Энсинк, Р., и ван Энгелен, Б. Г. (2009). Дисфагия присутствует, но незначительна при миотонической дистрофии 2 типа. Нервно-мышечная. Беспорядок. 19, 196–198. doi: 10.1016/j.nmd.2008.12.002
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Тилеман, А. А., Винке, А., ван Альфен, Н., ван Дейк, Дж. П., Пиллен, С., и ван Энгелен, Б. Г. (2012). Поражение скелетных мышц при миотонической дистрофии 2 типа. Сравнительное ультразвуковое исследование мышц. Нервно-мышечная. Беспорядок. 22, 492–499. doi: 10.1016/j.nmd.2012.01.006
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Уэдзуми, А., Фукада С., Ямамото Н., Такеда С. и Цучида К. (2010). Мезенхимальные предшественники, отличные от сателлитных клеток, способствуют образованию эктопических жировых клеток в скелетных мышцах. Нац. Клеточная биол. 12, 143–152. дои: 10.1038/ncb2014
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Урчуоло А., Кварта М., Морбидони В., Гаттаццо Ф., Молон С., Грумати П. и др. (2013). Коллаген VI регулирует самообновление сателлитных клеток и регенерацию мышц. Нац.коммун. 4, 1964. doi: 10.1038/ncomms2964
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Вальдес, М. Р., Ричардсон, Дж. А., Кляйн, У. Х., и Олсон, Е. Н. (2000). Неспособность Myf5 поддерживать миогенную дифференцировку без миогенина, MyoD и MRF4. Дев. биол. 219, 287–298. doi: 10.1006/dbio.2000.9621
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Виейра Н.М., Наславский М.С., Личинио Л., Кок Ф., Шлезингер Д., Вайнзоф, М., и соавт. (2014). Дефект РНК-процессирующего белка HNRPDL вызывает поясно-конечностную мышечную дистрофию 1G (LGMD1G). Гул. Мол. Жене. 23, 4103–4110. doi: 10.1093/hmg/ddu127
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Воханка С., Вытополь М., Беднарик Дж., Лукас З., Каданка З., Шильдбергер Дж. и соавт. (2001). Мутация в Х-сцепленном гене мышечной дистрофии Эмери-Дрейфуса у больного с проводниковой кардиомиопатией. Нервно-мышечная.Беспорядок. 11, 411–413. doi: 10.1016/S0960-8966(00)00206-6
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Вальц, П.К., Хиатт, К.К., Найду, М., и Халум, С.Л. (2008). Характеристика выживаемости и пролиферации стволовых клеток мышц гортани. Ларингоскоп 118, 1422–1426. doi: 10.1097/MLG.0b013e318173e188
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Вебстер, К., и Блау, Х.М. (1990). Ускоренное возрастное снижение репликативной продолжительности жизни миобластов мышечной дистрофии Дюшенна: значение для клеточной и генной терапии. Сомат. Ячейка Мол. Жене. 16, 557–565. дои: 10.1007/BF01233096
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Weintraub, H., Dwarki, VJ, Verma, I., Davis, R., Hollenberg, S., Snider, L., et al. (1991). Мышечно-специфическая активация транскрипции с помощью MyoD. Гены Дев. 5, 1377–1386. doi: 10.1101/gad.5.8.1377
Полнотекстовая перекрестная ссылка | Академия Google
Уайт, Р. Б., Биринкс, А. С., Ньокки, В. Ф., и Заммит, П. С.(2010). Динамика роста мышечных волокон в постнатальном развитии мышей. BMC Dev. биол. 10:21. дои: 10.1186/1471-213X-10-21
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Винокур, С.Т., Чен, Ю.В., Масни, П.С., Мартин, Дж.Х., Эмсен, Дж.Т., Тапскотт, С.Дж., и соавт. (2003). Профилирование экспрессии мышц FSHD подтверждает дефект на определенных стадиях миогенной дифференцировки. Гул. Мол. Жене. 12, 2895–2907. doi: 10.1093/hmg/ddg327
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Виртшафтер, Дж.Д., Феррингтон Д.А. и Маклун Л.К. (2004). Непрерывное ремоделирование экстраокулярных мышц взрослых как объяснение избирательной черепно-лицевой уязвимости при окулофарингеальной мышечной дистрофии. J. Нейроофтальмол. 24, 62–67. дои: 10.1097/00041327-200403000-00013
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ёсида А., Кобаяши К., Маня Х., Танигути К., Кано Х., Мизуно М. и др. (2001). Мышечная дистрофия и нарушение миграции нейронов, вызванные мутациями гликозилтрансферазы POMGnT1. Дев. Ячейка 1, 717–724. doi: 10.1016/S1534-5807(01)00070-3
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Зифко, У. А., Хан, А. Ф., Ремтулла, Х., Джордж, К. Ф., Вихлидал, В., и Болтон, К. Ф. (1996). Электрофизиологические исследования центрального и периферического дыхания при миотонической дистрофии. Мозг 119 (часть 6), 1911–1922 гг. doi: 10.1093/мозг/119.6.1911
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
квартир, квартиры в аренду в GK Palacio, Pune
Фильтры
Premium Filtersnew
апартаменты типа
1 RK
1 BHK
2 BHK
3 BHK
4 BHK
4+ BHK
Диапазон (в месяц)
Часто задаваемые вопросы
Могу ли я оплатить аренду дома с помощью кредитной карты через NoBroker? Если вы хотите оплатить аренду дома, депозит или даже символическую сумму, вы можете использовать NoBroker Pay.Это безопасная и зашифрованная платежная платформа. Если вы оплачиваете аренду с помощью кредитной карты, вы можете получать вознаграждения, кэшбэк и авиамили от вашего банка.Если у вас нет кредитной карты, вы можете оплатить аренду через NoBroker pay, используя свою дебетовую карту, наши счета UPI.
Какие еще есть населенные пункты рядом с ГК Паласио, Пуна, где можно снять квартиру? ГК Паласио окружен многими другими хорошо развитыми районами, такими как Моши, Чихали, Прадхикаран, Мои и Чимбали.Если вы не нашли хороший вариант в ГК Паласио, то вы всегда можете попробовать окрестности.Все они экономически ценены и имеют много, чтобы предложить вам.
Каковы лучшие учреждения в GK Palacio, Пуне и недвижимости в аренду поблизости?GK Palacio является популярным местным населением — если вы хотите найти дом в аренду в GK Palacio, проверьте этот список лучших вариантов предприятий.
Каковы виды недвижимости BHK в аренду в ГК Паласио, Пуна? В Пуне вы найдете много квартиры в аренду, которые имеют вариацию —1 РК — 1 комната и кухня в аренду в GK Palacio
1 BHK — 1 спальня, зал и кухня в аренду в GK Palacio
2 BHK — 2 спальни, зал и кухня в аренду GK Palacio
3 BHK — 3 спальни, зал и кухня в аренду в GK Palacio
4 BHK — 4 спальни, зал и кухня в аренду в GK Palacio
виллы — независимые дома в аренду в GK Palacio
Если вы живете один, то 1RK и 1BHK имеют самый смысл.Если вам нужно место для офиса дома, вы можете купить даже 2-комнатную квартиру.
Для тех, у кого есть семья, лучше выбрать 2-комнатную квартиру или более, в зависимости от количества человек в вашей семье.
В настоящее время большинство застройщиков имеют 2,5 BHK (квартиры с 2 с половиной спальнями) и так далее. Эти полукомнаты не такие большие, как обычная спальня, и вы можете использовать их как кабинет/домашний офис/кладовую и так далее.
Какие существуют типы мебели в квартире в аренду в Gk palacio, Пуна? Когда вы говорите о мебели для квартиры, есть три типа –Полностью меблирована
Частично меблированный
Без мебели
В полностью меблированной квартире в аренду в ГК Паласио, Пуна все доступно.От шкафов под телевизор до кроватей и кухонных шкафов и шкафов — в квартире будет все, что вам нужно.
Полу-меблированная квартира в аренду в Gk Palacio, Пуна будет иметь некоторую необходимую мебель, но не всю. Вы можете найти такие вещи, как встроенные шкафы, свет и вентиляторы, но вам нужно будет расставить другую мебель, которая вам нужна, например, стулья, столы и т. д.
В немеблированной квартире в аренду в ГК Паласио, Пуна, вам нужно будет расставить всю мебель самостоятельно.Это хороший вариант для тех людей, которые приобрели мебель и не хотят ее продавать. Если вы любите делать декор своими руками, то это лучший вариант для вас.
Как найти недвижимость в аренду без посредников в Gk palacio, Пуна?Лучший способ найти недвижимость в аренду без посредников в Gk palacio, Пуна — это выполнить поиск на NoBroker. В приложении и на веб-сайте NoBroker есть множество вариантов, которые вы можете выбрать. Самое приятное, что на сайте нет брокеров. Вы можете связаться НАПРЯМУЮ с арендодателями.
Какие еще услуги предоставляет NoBroker при аренде недвижимости в Gk palacio, Пуна? NoBroker учитывает все, что вам нужно при аренде дома в Пуне.Соглашений об аренде NoBroker, где вам не нужно будет посещать государственное учреждение, все будет сделано за вас и доставлено к вашему порогу.
Вы можете проверить NoBroker Pay для простых способов онлайн-оплаты аренды.
Упаковщики и грузчики NoBroker в Пуне для безопасной и надежной помощи при переезде.
NoBroker Home Services, где вы можете сделать уборку дома, нанять сантехников или электриков и даже плотников.
Мы можем предложить гораздо больше, просто посетите NoBroker для получения дополнительной информации.
Есть ли недвижимость в аренду для холостяков и семей в Gk palacio, Пуна?В Пуне есть много домов для холостяков и семей. Самый простой способ найти эти дома — на NoBroker. Когда вы получите страницу результатов поиска, просто используйте фильтры, чтобы выбрать дома, зарезервированные для холостяков и семей.Если он говорит и то, и другое, это означает, что арендодателю удобно иметь в собственности как холостяков, так и семьи.
Золотой Камуй Глава 190
Золотой Камуй Глава 190
Серьезно
** Спойлеры **
Название главы: 明日のために — Ради завтрашнего дня
Канто или ва яку сак но аранкеп синэп ка исам— Ничто не приходит с небес без целиСираши) попутно получили немного бессмертия Сугимото.
Наши завсегдатаи… просто особенные. Будь то Асирпа и ее ребята или Цуруми и компания. или Хиджиката и его веселая банда. И бродяга Огата. И фальшивая настоящая семья Чикапаси… И, конечно же, самые дорогие, оставшиеся дома — Хучи и Осома.
До сих пор единственными людьми, которые умирали, были заключенные, персонажи второстепенных арок и отцы всех форм и размеров.
Все когда-нибудь заканчивается. Как это зловещее путешествие.
Нода создал этих красивых сумасшедших людей…
Кироранке, несомненно, лучший боец и выживший наравне с Сугимото.
Я отдам Който. Он клевый.
Но Кироранке… это….было…
Оно
Взяло
Трое мужчин.
Три потрясающих солдата…
Убить одинокого тигра.
И он мог бы победить их даже в полуживом состоянии, если бы не тюлень, высунувший свою милую лысую голову.
Реакции:-
Кирорчан …
Шираиш милая крошка..
И Който, и Цукишима, и… Танигаки вернулся.
И бедный Асирпа. Это Кироранке Ниспа умирает кровавой ужасной смертью.
Она шепчет ему о воспоминаниях, но Който не слышит, так что, возможно, Цукишима тоже.
Глава прекрасен тем, что отправляет его. Это на самом деле душераздирающе. Это как бы заставляет почувствовать это чувство — знать, не зная, что «причина» не была ни тривиальной, ни важной. Что золото — это не друзья, которых вы приобрели на этом пути, и не само путешествие, но и не само настоящее золото.Это более глубоко, чем это, и все же, довольно просто. Что-то, что невозможно ни понять, ни осознать…
Киро, все кончено. Он принимает это. И он спокоен, зная, что ключ «отперт».
«Бесполезные» вещи, которые они делали в своем путешествии. Ох уж эти Стеньки и Бани!!
И София
Вероятно, она ему нравилась, даже когда она была худой.
И вот снова вопрос Асирпы остался без ответа.
Навсегда.
Подать жалобу потребителя на HRDOP, FINANCE TEAM DEPARTMENT LEVEL 14/190 QUEEN ST MELBOURNE AUSTRALIA
Сообщил: Картика
Контактная информация:
HRDOP, FINANCE TEAM DEPARTMENT LEVEL 14/190 QUEEN ST MELBOURNE AUSTRALIA Australia
G.К ИТ-решения
Привет, я Картика из Ченнаи
Я подала заявку на работу Тима через Интернет, и однажды мне позвонили с номера 9705665796, и она сказала, что ее зовут Джесси, и она дает работу Тима, когда я подал заявку онлайн и дал мне 10 ссылок в каких картинках где там какие я должен был изменить в pdf и так я должен был закончить за 10 дней и после этого когда я закончил с меня начали просить деньги типа я должен заплатить 671 рупий за фирменные зарядные устройства и затем позже попросили у меня 1700 рупий за минимальный баланс, а затем 7000 рупий, и я сказал нет, и мне позвонил человек, и он сказал, что он менеджер финансовой группы, поэтому он сказал, пришлите мне 7000, я дам вам зарплату через 5 минут и позже они снова попросили 1300.Раньше они отправляли обычные текстовые сообщения и присылали мне приветственное письмо и все такое, а потом они начали избегать моих сообщений, и все время, пока я пытался позвонить, оно было отключено. Прошло больше недели, я получил от них ответ, и я потерял все свои деньги, я потерял около 10500 рупий в секунду. Они сказали, что у них есть компания в Австралии, и заставили меня поверить. Пожалуйста, помогите мне вернуть мои деньги. Я потерял почти все. Они забрали все деньги, используя название вашей компании. Я действительно работал, верил им и в конце концов потерял все свои деньги
И номера, с которых я получил msg где – 9311821516
Пожалуйста, помогите мне вернуть мои деньги.
————————————————-
Как подать жалобу на HRDOP, FINANCE TEAM DEPARTMENT LEVEL 14/190 QUEEN ST MELBOURNE AUSTRALIA?
* Перейдите на страницу подачи жалобы
* Напишите HRDOP , FINANCE TEAM DEPARTMENT LEVEL 14/190 QUEEN ST MELBOURNE AUSTRALIA в разделе названия компании и подробно напишите свою жалобу
* Введите вашу личную информацию (адрес электронной почты не будет опубликован)
* Получите возмещение / замена / повреждения от HRDOP, УРОВЕНЬ ОТДЕЛА ФИНАНСОВОЙ КОМАНДЫ 14/190 QUEEN ST MELBOURNE AUSTRALIA.
|